Da in einem Referat vieles unklar blieb, möchte ich nun noch einmal versuchen, die Zusammenhänge in einfacheren Worten zu erklären:
Die Zelldifferenzierung (bzw. Genregulation) ist einer der wichtigsten Vorgänge, die in unserem Körper ablaufen. Fände keine Differenzierung statt, bestünden wir aus einer einzigen Sorte von Zellen. Da unser Körper aber täglich mit vielen verschiedenen Aufgaben konfrontiert wird, ist es sehr wichtig, dass eine "Arbeitsteilung" stattfinden kann. D.h., dass die vielen verschiedenen Zellen unseres Körpers unterschiedliche Aufgaben übernehmen. Das Zusammenspiel der einzelnen Zellarten macht uns erst zu dem, was wir sind.
Alle Zellen unseres Körpers, selbst die hochspezialisierten, gehen aber auf eine einzige Zelle zurück, nämlich die Zygote, die befruchtete Eizelle. Im Anschluss an die Befruchtung setzen zwei wichtige Prozesse ein, die Zellteilung und die Zelldifferenzierung. Obwohl so viele verschiedene Zellen entstehen, ist doch in allen Zellen unseres Körpers unser gesamtes Erbgut vorhanden (es gibt einige Ausnahmen, diese sollen jedoch nicht weiter beachtet werden). Dies konnte man durch das Experiment mit Dolly beweisen: Das Erbgut einer Euterzelle wird in den Ursprungszustand zurückversetzt, indem man es in eine Eizelle implantiert. Aus dieser Zelle kann sich nun wieder ein vollständiges Schaf entwickeln. Daraus konnte man schließen, dass auch in einer Euterzelle die Information für ein komplettes Schaf vorhanden ist. Die Gene, die für andere Gewebearten stehen, müssen also ausgeschalten (inaktiv) sein, während die Gene, die für Eutergewebe stehen, angeschalten (aktiv) sind. Wie die Aktivierung bzw. die Inaktivierung abläuft ist erst ansatzweise erforscht und verstanden. Trotzdem gibt es einige Hiweise, die für die Gewebezucht äußerst wichtig sind.
Zellen, die dem Körper entnommen und im Reagenzglas zur Teilung angeregt werden, verlieren einige ihrer typischen Eigenschaften, d.h., die sogenannte Dedifferenzierung setzt ein. Ursache für die Dedifferenzierung sind vermutlich die erhöhte Teilungsrate einerseits und der Verlust der gewebetypischen Umwelt andererseits (auf die genauen Zusammenhänge werde ich später zurückkommen). Will man aus diesen Zellen, die nun wieder in ihren Ursprungszustand zurückversetzt sind, eine bestimmte Gewebeart züchten, muss man die Natur nachahmen. Dafür studiert man die Vorgänge, die im Körper ablaufen und zur Differenzierung der Zellen führen, und wendet sie im Reagenzglas an. Diese Vorgänge möchte ich im Folgenden näher erklären.
![]()
In unseren Zellen sind zwei wesentliche Stoffe vorhanden: Chromatin und Proteine. Chromatin besteht aus DNA und Proteinen. Proteine sind Ketten aus Aminosäuren. Unsere Gene liegen auf der DNA. Der Begriff Gen bezeichnet einen Abschnitt auf der DNA, also eine Sequenz (Abfolge) der vier Basen Adenin, Thymin, Guanin und Cytosin. Das Gen allein zeigt aber noch keine Wirkung in unserem Körper. Es muss zuerst in ein Protein übersetzt werden. Jedes Gen steht für ein Protein und die Proteine sind die Bausteine unseres Körpers. Daraus wird ersichtlich, dass Gene in Proteine übersetzt werden müssen. Wird diese Übersetzung an einer Stelle unterbrochen, kommt das entsprechende Gen nicht zur Wirkung. Diese Unterbrechung hat also dieselben Folgen, die ein Nicht-Vorhandensein des Gens auch hätte.
Die Übersetzung vom Gen ins Protein erfolgt in zwei Schritten. An erster Stelle steht die Transkription: DNA wird in mRNA (Boten-RNA) umgeschrieben. mRNA ist einsträngig und kann im Gegensatz zu DNA den Zellkern verlassen. Außerdem weist sie einige weitere Besonderheiten auf, die ich hier aber nicht ausführlicher beschreiben möchte. An zweiter Stelle steht die Translation: Die mRNA verlässt den Zellkern. Die Ribosomen, kleine Zellorganellen, die aus zwei Untereinheiten bestehen, lagern sich an die mRNA an. Nun wird t-RNA (transfer-RNA) an die m-RNA gebunden. t-RNA ist Träger der Aminosäuren. Jeweils drei Basen auf der m-RNA bilden die "Andockstelle" für einen Komplex aus t-RNA und einer Aminosäure. Die Aneinanderreihung der Aminosäuren ergibt das Protein. Sobald entweder die Transkription oder die Translation verhindert wird, wird auch die Bildung des Proteins verhindert.
![]()
Transkription:
Zum Transkriptionsstart werden neben RNA-Polymerase, einem Enzym, das die Bildung von m-RNA bewirkt, auch einige Transkriptionsfaktoren (bestimmt Proteine) benötigt. Diese Transkriptionsfaktoren binden an der TATA-Box, einer kurzen Sequenz vor dem Gen, das transkribiert werden soll. Gemeinsam bilden TATA-Box und Transkriptionsfaktoren den sogenannten Initiationskomplex, der die Startstelle für die Transkription bildet (hier beginnt die RNA-Polymerase, das Gen, also die DNA, in mRNA zu übersetzen). Bei den hier erwähnten Transkriptionsfaktoren handelt es sich um Proteine, die für den Transkriptionsstart benötigt werden. Es gibt aber auch noch andere Transkriptionsfaktoren. Diese binden nicht an der TATA-Box, sondern an anderen kurzen Sequenzen auf der DNA, den sogenannten Regulatorsequenzen. Wie der Name schon sagt, haben diese Sequenzen eine regulierende Auswirkung auf die Transkription, d.h., sie können die Transkription entweder verhindern oder verstärken. Die Sequenzen, die die Transkription verhindern, nennt man Silencer. Die Sequenzen, die die Transkription verstärken, bezeichnet man als Promotor, bzw. als Enhancer. Der einzige Unterschied zwischen Promotor und Enhancer besteht darin, dass Enhancer sehr viel weiter von dem Gen, das transkribiert werden soll, entfernt liegen. Durch Schleifenbildung der DNA gelangen sie jedoch in unmittelbare Nähe des Gens. Enhancer können vor oder hinter dem Gen liegen.
![]()
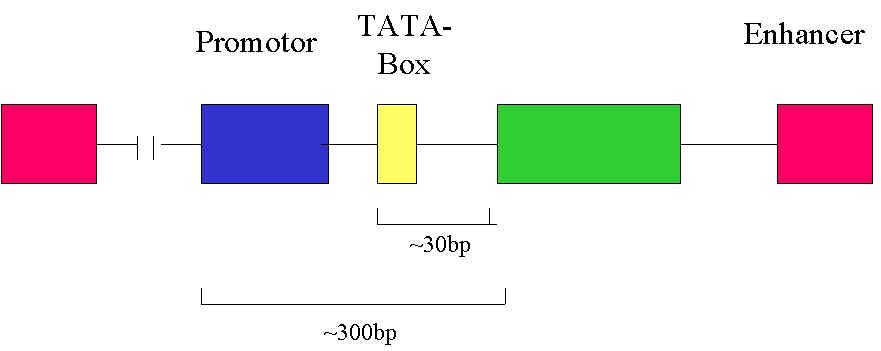
In der Abbildung sind die Enhancer, die vor oder hinter dem Gen liegen können, rot dargestellt, der Promotor blau, die TATA-Box gelb und das Gen, das transkribiert werden soll grün. Die TATA-Box bildet gemeinsam mit Proteinen, den Transkriptionsfaktoren, den Startpunkt für die RNA-Polymerase. Enhancer und Promotor bilden Bindungsstellen für weitere Proteine (andere Transkriptionsfaktoren), die die Transkription verstärken. Silencer sind in der Abbildung nicht dargestellt.
Die Regulatorsequenzen haben selbstverständlich nur dann Auswirkungen auf die Transkription, wenn Proteine an sie gebunden sind.
Diese DNA-bindenden Proteine werden in drei Gruppen eingeteilt, die ich kurz charakterisieren möchte:
![]()
1.) Helix-Knick-Helix-Motiv
Die Proteine dieser Gruppe binden als Dimere an die DNA (Dimere sind Verbindungen aus zwei gleichartigen Molekülteilen. Die Proteine, die Dimere bilden, sind also als einzelne Moleküle nicht aktiv, sondern nur wenn beide Untereinheiten eine Einheit bilden. Vergleichen könnte man dieses Prinzip mit einem Druckknopf, der seine Funktion auch erst erfüllt, wenn beide Teile zusammengesteckt werden.). Weiterhin besitzen sie drei alpha-helikale-Abschnitte (eine alpha-Helix besitzt eine Raumstruktur von Kettenmolekülen mit einer rechtsgängigen spiralförmigen Windung um eine Zentralachse). Diese Abschnitte sind durch kurze Schleifen voneinander getrennt. Eine Helix jeder Dimerenuntereinheit besetzt die weite Furche der DNA über zwei Windungen hinweg (die DNA hat die Form einer Doppelhelix und weist abwechselnd eine weite und eine enge Furche auf).
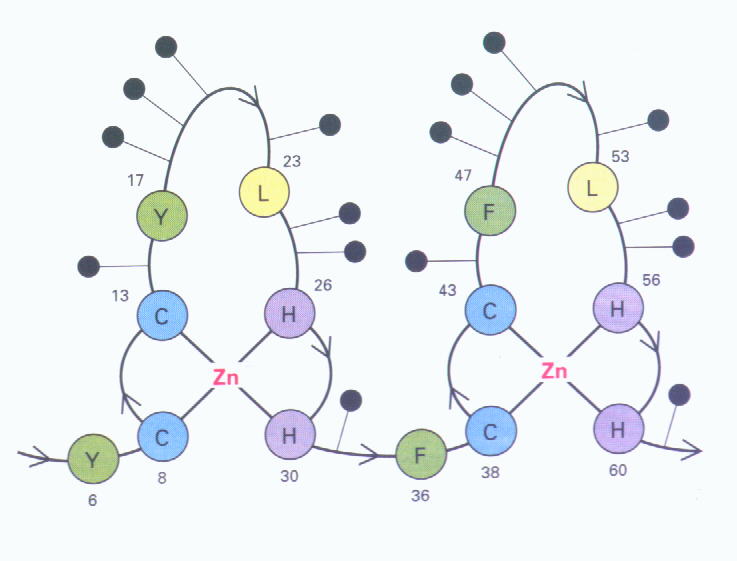
2.) Zinkfinger-Proteine
Diese Proteine haben neun repetitive Domänen, die Cystein und Histidin (Aminosäuren) enthalten (als Domänen bezeichnet man in einer kompakten Struktur gefaltete Teilstücke eines Proteins; als repetitiv bezeichnet man die Eigenschaft von Sequenzen, in zahlreichen Kopien vorzukommen). Zink wird in den meisten Fällen an Zystein und Histidin gebunden. In der Zinkfingerstruktur bilden die sich wiederholenden Domänen Schleifen, so dass das Zinkion von einem Cystein- und einem Histidinpaar fixiert werden kann.
![]()
3.) Amphiphile helikale Proteine (Helix-Schleife-Helix-Proteine):
Ein 100-200 Aminosäuren langer Abschnitt am C-Terminus dieser Proteine kann sich falten, wobei er zwei amphiphile alpha-Helices bildet (amphiphil bedeutet, dass ein hydrophiles und ein hydrophobes Ende vorhanden sind). Auch diese Proteine bilden Dimere. Die helikalen Abschnitte werden zur Dimerisierung benötigt.
Dimeres Helix-Schleife-Helix-Protein: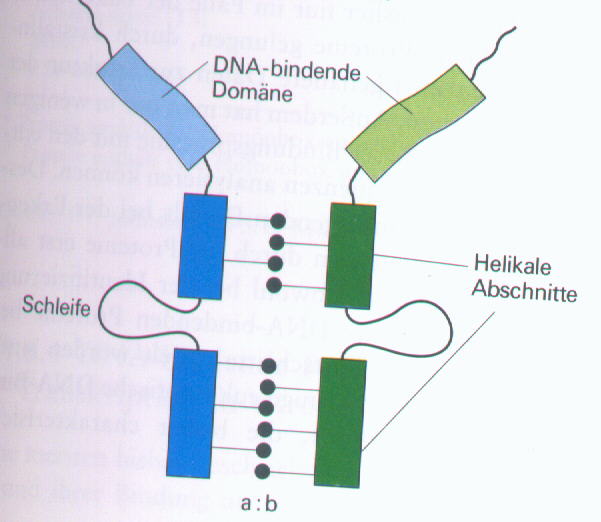
![]()
Die Auswirkung dieser Proteine auf die DNA kann man sich so vorstellen:
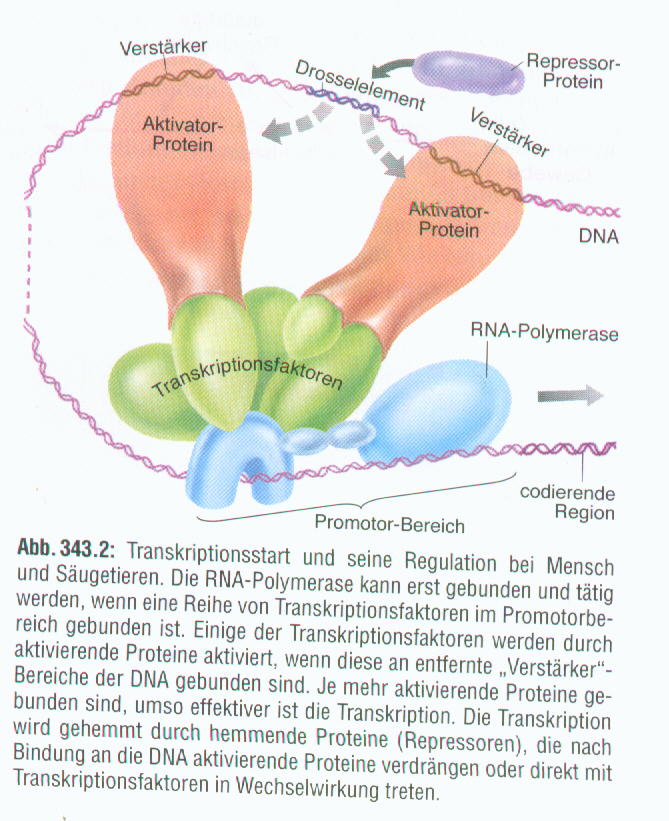
Offensichtlich ist, dass die Proteine nicht nur Auswirkungen auf die Transkriptionsaktivität haben, sondern auch auf die DNA-Struktur. So konnte man beobachten, dass im Bereich der inaktiven Gene die DNA stark kondensiert. Die Transkription findet während der Interphase statt, also zu einem Zeitpunkt, zu dem sich die Zelle nicht teilt. Die DNA liegt als lockere Struktur im Kern, während sie bei der Mitose auf Proteine aufgewickelt und so stark verkürzt (kondensiert) wird. Doch auch während der Interphase sind einige Stellen der DNA stark kondensiert. An diesen Stellen kann keine mRNA nachgewiesen werden, folglich findet auch keine Transkription statt. Die Transkriptionsfaktoren, die dieTranskription hemmen, führen also gleichzeitig zur Kondensation der DNA in dem Bereich, in dem sie wirken.
Bisher habe ich noch nicht darüber gesprochen, wie Proteine dazu gebracht werden, an die Regulatorsequenzen zu binden.
Die Aktivierung der Proteine ist abhängig von Signalen. Signale sind beispielsweise Hormone oder Wachstumsfaktoren.
Steoridhormone:
Steoridhormone können durch Diffusion durch die lipophile Biomembran in die Zelle gelangen. In der Zelle sind Rezeptoren vorhanden, die an die Regulatorsequenzen auf der DNA binden können. Diese Rezeptoren sind inaktiv. Erst wenn das Hormon daran gebunden wird, ist der Komplex aus Rezeptor und Hormon aktiv. Der Komplex gelangt in den Kern und bindet sich an ein Response-Element (kurze Sequenz innerhalb der Regulatorsequenzen, an die sich Proteine binden können).
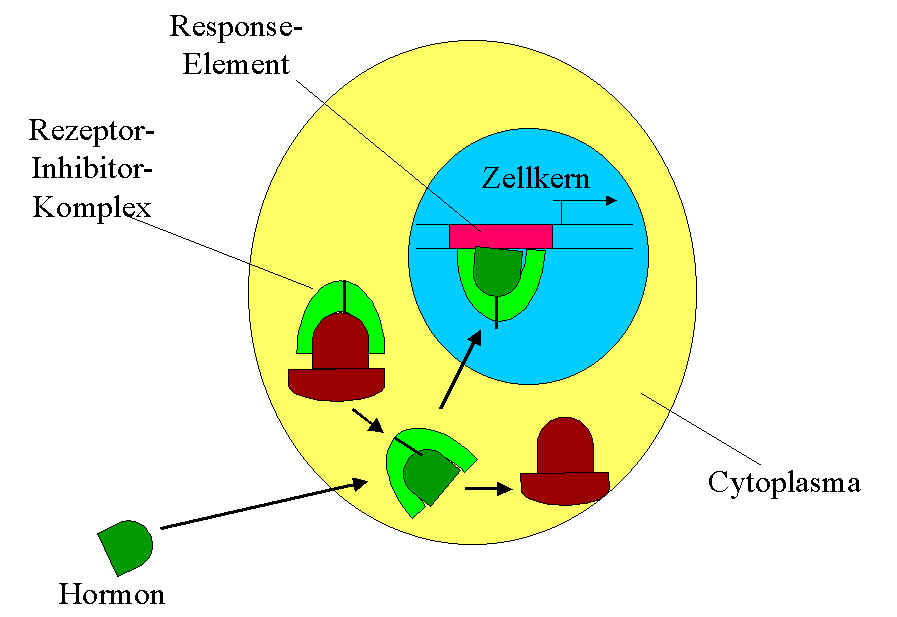
Das Hormon gelangt in die Zelle und bindet an den Rezeptor, der noch durch einen Inhibitor blockiert ist. Gemeinsam werden Rezeptor (hellgrün) und Hormon (dunkelgrün) am Response-Element gebunden.
Rezeptor-Ligand-Wechselwirkungen auf der Zelloberfläche
Auch auf der Zelloberfläche befinden sich Rezeptoren, an die Stoffe gebunden werden können. Die Wechselwirkungen, die aus diesen Bindungen resultieren, bewirken einen Anstieg der Second-Messenger-Moleküle in der Zelle. Dabei handelt es sich um bestimmte Moleküle wie Ca++, Diacylglycerin oder cyclische Nukleotide. Diese Moleküle bewirken die Aktivierung von Proteinkinasen. Dies sind Enzyme, die die Phosphorylierung von Molekülen bewirken. Diese Phosphorylierung bewirkt wiederum eine Aktivierung der betroffenen Proteine.
Diese Signalwirkungen versucht man im Reagenzglas bei der Gewebezucht nachzuahmen, indem man beispielsweise Hormone oder Wachstumsfaktoren hinzugibt. Bisher gelingt es allerdings nur, jeweils eine Art von Zellen zu züchten. Man schafft es aber auch schon, in diesen Geweben gefäßartige Strukturen zu züchten, doch diese Versuche befinden sich noch weitgehend in einem Anfangsstadium.
![]()
Translation:
Die Translation kann verhindert werden, wenn die mRNA zu langsam in das Cytoplasma gelangt oder wenn ihre Abbaugeschwindigkeit zu hoch ist. Wird die mRNA abgebaut bevor sie in Aminosäuren übersetzt werden kann, kommt auch das entsprechende Gen nicht zur Wirkung.
Eine Realisierungschance dieser Zusammenhänge in der Gewebezucht besteht mit Sicherheit, denn schon jetzt werden die hier dargelegten Forschungsergebnisse angewendet. Das Hauptproblem besteht allerdings darin, dass man noch zu wenig über die Differenzierung einerseits und die Kommunikation zwischen den verschiedenen Zellarten andererseits weiß. Wenn man komplexe Organe schaffen will, ist dieses Wissen aber unerläßlich. Deshalb wird es wohl noch länger dauern, bis im Labor das erste Herz entsteht.
Literatur:
DARNELL, James / LODISH, Harvey / BALTIMORE, David: Molekulare Zellbiologie
HAGEMANN, Rudolf: Allgemeine Genetik
BAYRHUBER, Horst / KULL, Ulrich: Linder Biologie
Lexikon Medizin
Bilder:
DARNELL, James / LODISH, Harvey / BALTIMORE, David: Molekulare Zellbiologie
BAYRHUBER, Horst / KULL, Ulrich: Linder Biologie
Fragen und Kommentare an Simone Wiesler © Faust-Gymnasium Staufen, letztes update 25.01. 2001