- Die
Sorge um die Menschen und ihr Schicksal muss stets das
Hauptinteresse allen technischen Strebens bilden, die großen
ungelösten Fragen der Organisation der Arbeit und der
Güterverteilung, damit die Erzeugnisse unseres Geistes den
Menschengeschlecht zum Segen gereichen und nicht zum Fluche.
Vergesst dies nie über Euren Zeichnungen und Gleichungen.
- Einstein

|
-
- Tissue
Engineering
- Inhalt
-
- Klicken Sie auf einen
Gliederungspunkt!
-
-
-
- Anhang
-
- a) Glossar
- b) Literaturliste
- c) Abbildungsnachweis
- d) weiterführend kommentierte
Bibliographie
-
-
-
-
- Der
Inhalt meiner Seminararbeit lässt sich eigentlich in wenigen Sätzen
zusammenfassen: Durch bestimmte Faktoren werden auf molekulargenetischer
Ebene Vorgänge ausgelöst, die in vivo die Zelldifferenzierung
verursachen. Diese Vorgänge werden in vitro nachgeahmt, um Gewebe zu züchten,
das, wenn es in den Körper implantiert wird, eine bestimmte Funktion übernehmen
kann.
- Genauer
betrachtet ist dieser Vorgang natürlich sehr viel komplizierter. Meiner
Ansicht nach handelt es sich jedoch gerade wegen dieser Komplexität bei
Tissue Engineering um eine der interessantesten und vielversprechendsten
Techniken auf dem Gebiet der Biotechnologie und es lohnt sich durchaus,
sich genauer damit zu beschäftigen.
- Warum
eine der vielversprechendsten Techniken? In den USA entstehen „ca. 400
Mrd. $ jährliche Kosten durch Organ und Gewebeschäden“[1].
Leider gibt es viel weniger Organe, als benötigt werden, und wenn ein
Patient ein Spenderorgan bekommt, können Abstoßungsreaktionen
auftreten. Die Gefahren einer Transplantation sind deshalb immer sehr
hoch. Die moderne Gewebezucht versucht, Gewebe und Organe herzustellen,
deren Zellen mit körpereigenen identisch sind. Dadurch sollen
Immunreaktionen vermieden werden.
- Zudem
ergeben sich neue Möglichkeiten für die Alzheimer- und die
Parkinsonforschung. Durch Züchtung von Nervenzellen will man zerstörte
Hirnregionen wieder herstellen und den Patienten ein besseres Leben ermöglichen.
- In
meiner schriftlichen Arbeit möchte ich mich damit befassen, wie man
Organe und Gewebe züchtet, welche molekulargenetischen Prozesse
ausgenutzt werden können bzw. beachtet werden müssen und was auf
diesem Gebiet bisher erreicht wurde.
- Meine Arbeit
kann insgesamt in drei Teile gegliedert werden: Im ersten Teil möchte
ich mich mit den molekulargenetischen Grundlagen der Zelldifferenzierung
beschäftigen, im zweiten Teil damit, wie man diese Prozesse in vitro
nachzuahmen versucht, um funktionelles Gewebe zu erhalten. Diese beiden
Teile sollen einerseits eine Beschreibung dessen liefern, was sich tatsächlich
im Organismus eines Lebewesens abspielt. Denn die natürlich ablaufenden
Vorgänge müssen genau beachtet werden. Andererseits möchte ich
beschreiben, wie man in vitro beobachtet, wie lebende Zellen von
verschiedenen Substanzen beeinflusst werden und wie die Ausbildung
unterschiedlicher Merkmale begünstigt wird. Dabei sollte man nie
vergessen, dass die Situation für entstehendes Gewebe in vitro anders
ist als in vivo.
- Da man, was
diese Beobachtungen angeht, noch relativ am Anfang steht, können die
dargestellten Methoden und Vorgänge nicht vollständig beschrieben
sein. Ich kann daher nur darüber schreiben, was bisher erforscht wurde
und muss dabei sämtliche Spekulationen in Bezug auf die Zukunft
beiseite lassen.
- Der dritte
Teil enthält die Auswertung dessen, was ich in den beiden vorangehenden
Kapiteln erarbeitet habe und eine Beleuchtung des Themas unter
gesellschaftlichen, ethischen und sozialen Aspekten.
-
- 2. Zelldifferenzierung
-
- 2.1. in vivo
-
- Wie es zur
Differenzierung kommt, ist bisher nur ansatzweise erforscht und
verstanden.
- Die
bisherigen Forschungsergebnisse auf diesem Gebiet möchte ich nun erläutern,
um anschließend darauf eingehen zu können, wie sich der Mensch diese
Erkenntnisse zunutze machen kann, um selbst in vitro die Differenzierung
auszulösen und spezialisierte Zellen zu erhalten.
-
- 2.1.1.
Bedeutung und Beginn der Zelldifferenzierung
-
- Prokaryonten
sind Einzeller. In einer einzigen Zelle laufen sämtliche
lebensnotwendigen Vorgänge ab. Im Laufe der Evolution sind aus diesen
Einzellern Vielzeller, Eukaryonten, entstanden, die zum Teil aus einer
unvorstellbar großen Zahl von Zellen bestehen.
- Nehmen wir als Beispiel den
Menschen: Er besteht aus 6x1013
Zellen.
Diese Zellen sind wesentlich komplizierter gebaut als die Protozyten.
Zudem ist nicht jede einzelne von ihnen für jeden im Körper
ablaufenden Vorgang zuständig.
- Eine
Hautzelle hat beispielsweise völlig andere Aufgaben als eine
Nervenzelle; ebensowenig kann eine Muskelzelle die Aufgabe einer
Darmzelle erfüllen oder eine Leberzelle die Aufgaben einer Blutzelle.
- Im Organismus
wären schwere Krankheiten die Folge, wenn sich beispielsweise in der
Leber plötzlich Hautzellen ansiedeln würden. An diesem Beispiel lässt
sich erkennen, wie wichtig es ist, dass verschiedene Zellen jeweils an
dem Ort bleiben, wo sie hingehören.
- Doch warum
besteht der menschliche Körper überhaupt aus so vielen verschiedenen
Zellen? Warum kann nicht – wie bei Prokaryonten – eine einzige
Zellart sämtliche Aufgaben übernehmen? Die Antwort liegt in der
Komplexität des menschlichen Organismus. Wir sind sehr viel höher
organisierte Wesen, als Bakterien. Mit entsprechend mehr
Herausforderungen muss unser Körper fertig werden. Dass er dies tatsächlich
kann, ist die Folge komplizierter Prozesse, durch die hochspezialisierte
Zellen entstehen.
- Letztendlich
gehen jedoch alle Zellen eines Organismus auf eine einzige Zygote zurück.
Durch mitotische Teilungen entstehen aus dieser Zygote Tochterzellen,
die bis zum Dreizellstadium totipotent sind, d.h., sie sind noch nicht
differenziert und besitzen die Fähigkeit zu jeder beliebigen Somazelle
zu werden.
Erst danach beginnen die Zellen sich zu spezialisieren. Einige dieser
Differenzierungen können unter bestimmten Voraussetzungen wieder rückgängig
gemacht werden, da sie nicht irreversibel sind.
-
-
- 2.1.2.
Strukturgene, Homöobox- und Regulatorgene
-
- Verantwortlich
für die Differenzierung sind Prozesse, die die Proteinbiosynthese
unterbrechen oder verstärken. Dies geschieht durch eine Erhöhung bzw.
Verminderung der Transkriptionsaktivität oder das Verhindern der
Translation. Im Folgenden möchte ich näher auf die
Transkriptionsaktivität eingehen. Neben den sogenannten Strukturgenen,
die jeweils für ein Protein codieren, gibt es auf der DNA auch lange,
scheinbar funktionslose, Abschnitte, die häufig „regulatorische
Aufgaben“[4]
haben. Des weiteren gibt es auch sogenannte Homöoboxgene. Sie besitzen
konservierte DNA-Abschnitte, die aus 183 Basenpaaren (Homöobox)
bestehen; das entstehende Proteinsegment besteht also aus 61 Aminosäuren.
Man bezeichnet es als Helix-Knick-Helix-Motiv (Helix-turn-helix-Motiv).[5]
Diese, sowie die Produkte einiger weiterer Gengruppen, können
spezifisch an Regulatorsequenzen binden und den Transkriptionsablauf
beeinflussen. Man bezeichnet sie deshalb als Transkriptionsfaktoren. Es
werden verschiedene Arten von Regulatorsequenzen unterschieden: Promotor
und Enhancer verstärken die Transkriptionsaktivität, während Silencer
diese Aktivität vermindern oder sogar völlig verhindern.[6] Enhancer liegen häufig sehr weit (bis zu 50kb) vom zu
transkribierenden Gen entfernt. Sie liegen meistens stromaufwärts,
seltener stromabwärts von diesem Gen. Promotoren liegen sehr viel näher
am Gen. Der einzige Unterschied zwischen diesen beiden
Regulatorsequenzen besteht also in ihrer Entfernung. Durch
Schleifenbildung gelangen Enhancer in unmittelbare Nähe des zu
transkribierenden Gens.[7]
Alle diese Regulatorsequenzen sind Bindungsstellen für
Transkriptionsfaktoren.
-
-
- 2.1.3.Charakterisierung
der Transkriptionsfaktoren
-
- Zum Transkriptionsstart werden
Proteine benötigt, die ebenso wie die Proteine, die regulierend auf die
Transkription wirken, Transkriptionsfaktoren genannt werden.
Bindungsstelle für Transkriptionsfaktoren, die zum Transkriptionsstart
benötigt werden, wie der TATA-Faktor, ist die sogenannte TATA-Box, die
ca. 30 Nukleotide stromaufwärts von der RNA-Startstelle entfernt liegt.
- Transkriptionsfaktoren, die
regulierende Funktion haben, binden an Silencer, Promotor oder Enhancer;
sie können durch ihre Struktur unterschieden und in drei Gruppen
eingeteilt werden. Man bezeichnet sie als Helix-Knick-Helix-Motiv,
Zinkfinger oder amphiphiles Helix-Motiv.
- Helix-Knick-Helix-Motiv:
- Die
Bezeichnung haben diese Proteine von drei α-helikalen Abschnitten,
die durch Schleifen voneinander getrennt sind. An der DNA binden sie als
Dimere.
Eine Proteinhelix besetzt die weite Furche der DNA Helix. Sie steht dort
mit den Basenpaaren in Kontakt. Über dieser Helix liegt eine zweite und
nimmt mit „anderen Proteinen des Transkriptionsapparates“
Verbindung auf.
- Die Homöobox
Sequenzen sind bei vielen Lebewesen homolog. Sowohl in der Helix 2 als
auch in der Helix 3 findet man an Schlüsselpositionen Aminosäuren, die
sich chemisch ähneln.
-
- Abb.1: 1,2,3: alpha-helikale Abschnitte
-
- Zinkfinger:
- Ihren
Namen haben diese Proteine von ihren charakteristischen Schleifen, die
von neun repetitiven Domänen gebildet werden, und einem Zinkion. Die
sich wiederholenden Abschnitte enthalten sowohl Cystein als auch
Histidin, Aminosäuren, die Zink binden können. Die Schleifenbildung
ermöglicht, dass jeweils ein Cystein-und ein Histidinpaar ein Zinkion
binden. Wichtig für die DNA-Bindung sind Phenylalanin- Thyrosin und
Leucinreste in den Schleifen [12].
-
-
- Abb.
2:C:
Cystein; H: Histidin; Zn: Zink
-
- Amphiphiles
Helix-Motiv:
- Diese
Proteingruppe kann wiederum in zwei Untergruppen aufgeteilt werden.
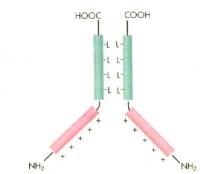
- Leucinreißverschluss-Proteine
besitzen vier oder fünf Leucin-Bausteine, die jeweils im Abstand von
sieben Aminosäuren auftreten. Diese Domänen sind hydrophob und ermöglichen
die Bildung von Dimeren, indem zwei derartige Proteine miteinander in
Kontakt treten. Die andere Seite dieser Proteine enthält die positiv
geladenen Aminosäuren Arginin und Lysin. Mit dieser Domäne kann das
Protein an die DNA binden.
-
-
- Abb.
3:
L: Leucin (Aminosäure); +: positive Ladung
-
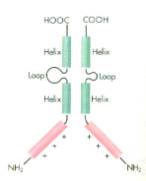
- Helix-Schleife-Helix-Proteine
(helix-loop-helix)
binden ebenfalls mit einer positiv geladenen Domäne als Dimere an die
DNA[13].
Eine 100-200 Aminosäuren lange Sequenz faltet sich und bildet dabei
zwei amphiphile α-Helices. Die benachbarte Sequenz enthält
besonders viele basische Aminosäuren[14].
-
-
- Abb.
4:
Loop: Schleife
-
- Wesentlich ist, dass alle
Proteine, die ich hier beschrieben habe, jeweils einen Abschnitt haben,
der an die DNA binden kann und einen, der regulierend auf die
Transkription wirkt.
-
-
- 2.1.4. Aktivierung der
Transkriptionsfaktoren
-
- Transkriptionsfaktoren
liegen meist als inaktive Komplexe im Cytoplasma vor. Damit sie an die
Regulatorsequenzen gebunden werden, müssen sie zuerst aktiviert werden.
Die Aktivierung erfolgt durch eine Reihe von Steuerungssubstanzen, die
offenbar in unterschiedlicher Konzentration in den verschiedenen Geweben
auftreten und deshalb die Spezialisierung ermöglichen. Einige dieser
Steuerungsfaktoren möchte ich im Folgenden näher beschreiben.
- Steoridhormone:
- Steoridhormone
sind lipophil und können deshalb durch die Biomembran der Zellen
diffundieren und ins Plasma gelangen. In der Zelle können sie an
Rezeptoren binden, die sich spezifisch an Response-Elemente heften und
somit aktive Transkriptionsfaktoren sind. Diese Rezeptoren heften sich
nur an die DNA, wenn sie vorher ein Steroidhormon gebunden haben, da sie
Teil eines großen Proteinkomplexes sind. Dadurch werden sie daran
gehindert, in den Kern zu gelangen.
- Das Hormon
bewirkt, dass der Rezeptor aus dem Komplex gelöst wird und nun in den
Kern gelangen kann, wo er mit der DNA eine Bindung eingeht.
- Rezeptor-Ligand-Wechselwirkungen:
- Neben den Steuerungssubstanzen,
die wie die Steroidhormone direkt bewirken, dass ein Protein aktiviert
und an die DNA gebunden wird, gibt es auch solche, die durch
Wechselwirkungen die Freisetzung weiterer Moleküle bewirken, die
wiederum Einfluss auf die Genregulation haben. Auch auf der Zelloberfläche
gibt es Rezeptoren, an die sich Stoffe wie z.B. α-Interferon
binden, die nicht wie Steroidhormone durch die Membran diffundieren können.
Auf der Zelloberfläche kommt es zu Wechselwirkungen und die
Konzentration der sogenannten second-messenger-Moleküle steigt an. Ein
besonders wichtiges second-messenger-Molekül ist das zyklische
Adenosin-3`,5`-monophosphat (cAMP), das die Proteinkinase A (PKA)
aktiviert. PKA ist in der Lage, bestimmte Proteine zu phosphorylieren
und damit zu aktivieren. Im Gegensatz zur Aktivierung, die durch
Steroidhormone erfolgt, wird jedoch nicht die DNA-Bindungsdomäne
aktiviert, sondern die Domäne, die auf den Transkriptionskomplex wirkt.
Weitere second-messenger-Moleküle, die eine Freisetzung von
Proteinkinasen bewirken, sind cyclisches Guanosin 3`,5`-monophosphat (cGMP),
Ca++ und Diacylglycerin.
- Die
Signalmoleküle, die an Rezeptoren auf der Zelloberfläche binden, sind
z.B. das Protein Sonic hedgehog (Shh), sowie Wachstumsfaktoren (z.B.:
Plateletderived growth factor, Fibroblastenwachstumsfaktoren, epidermale
Wachstumsfaktoren, insulinähnliche Wachstumsfaktoren, etc.).
-
-
- 2.2. in vitro
-
- Bringt
man Zellen außerhalb des Körpers in ein Nährmedium, laufen andere
Vorgänge ab, als in ihrem natürlichen Umfeld. Dies liegt vor allem
daran, dass Signalsubstanzen, die zur Aktivierung von
Transkriptionsfaktoren und somit zur Expression der Charakteristika
einer Zelle führen, nicht in derselben Konzentration im Nährmedium
auftreten wie im jeweiligen Gewebe.
-
-
- 2.2.1. Dedifferenzierung
-
- Die Dedifferenzierung der Zellen
ist ein Phänomen, das in vitro auftritt. Die Zellen verlieren ihre
ursprünglich charakteristischen Merkmale und werden scheinbar
reembryonalisiert. Dies ist offenbar bei allen Körperzellen der Fall
zu. Ein berühmtes Beispiel dafür ist das Schaf Dolly, das aus einer
Euterzelle geklont wurde, deren Erbgut in eine Spendereizelle
eingebracht wurde. Das Erbgut wurde daraufhin in den Ursprungszustand
zurückversetzt.
-
-
- 2.2.2. Gründe für die Dedifferenzierung
-
- Schuld an der
Dedifferenzierung sind vor allem die mitotischen Teilungen, die in vitro
provoziert werden. Eine Differenzierung findet immer während der
Interphase statt, denn während der Mitose werden die Chromosomen stark
kondensiert. Sie können deshalb an keiner Stelle entspiralisiert und
somit auch nicht transkribiert werden. Je häufiger sich die Zellen
teilen, desto mehr verlieren sie ihr ursprüngliches Aussehen. Dieses
erlangen sie in vitro auch nicht wieder, da Signalmoleküle zur
Aktivierung von Transkriptionsfaktoren nicht in der idealen
Konzentration vorhanden sind.
-
-
- 2.2.3. Probleme in Bezug auf
Tissue Engineering
-
- Im Fall von
Dolly war eine Dedifferenzierung durchaus von Vorteil. Weniger erwünscht
ist sie in der Gewebezucht, da man ja hier eine Vielzahl von
hochspezialisierten Zellen erhalten möchte. Es sind also genaue
Messungen der Faktoren, die im Gewebe vorkommen, erforderlich, um die
Natur nachzuahmen und die gewünschte Differenzierung zu erhalten. Genau
dies versucht man bei Tissue Engineering.
-
-
-
- 3.
Tissue Engineering
-
-
- 3.1.
Wozu braucht man künstliches Gewebe?
-
- Schlagzeilen
wie „Organtransplantation geht neue Wege“ (SZ, 10.2.1999) oder
„Haut aus der Retorte“ (SZ 9.3.1999) weisen darauf hin: Die
Nachfrage nach Organen und verschiedenen Gewebearten ist sehr groß und
kann mit den herkömmlichen Methoden nur unzureichend befriedigt werden.
„In den USA entstehen ca. 400 Mrd $ jährliche Kosten durch Organ- und
Gewebeschäden”
– auch dies zeigt, dass es wichtig ist, neue Methoden zu entwickeln,
Menschen mit derartig schweren Schäden zu helfen. Die Anwendungsgebiete
sind vielfältig. Man benötigt neue Herzklappen, Darmzellen, Knorpel-
und Knochenzellen. Das bekannteste Beispiel für ein Anwendungsgebiet
des Tissue Engineering ist die „Haut aus der Tube“ für Menschen die
z.B. an schweren Verbrennungen leiden.
- Es scheinen
sich jedoch auch weitere Möglichkeiten aufzutun. Die Neubildung jener
Nervenzellen, die bei Parkinson im Gehirn absterben,
verspricht Heilungschancen für diese Krankheit.
- Noch ist es
sehr schwierig, das gewünschte Gewebe zu züchten. Die
Dedifferenzierung, die ich bereits im vorhergehenden Kapitel beschrieben
habe, ist ein Grund dafür. Die Bedingungen für die Entwicklung von
funktionellen Geweben unterscheiden sich in vitro sehr stark von den
Bedingungen in vivo. Deshalb versucht man nun, auch in vitro Situationen
zu schaffen, die denen im lebenden Gewebe möglichst ähnlich sind.
-
-
- 3.2.1.
Materialien und Geräte
-
- Funktionelles
Gewebe kann nur in gewebetypischer Umwelt hergestellt werden.
- Es ist darauf
zu achten, dass die Herstellung von Gewebe in drei Schritten ablaufen
muss: Zuerst muss man dafür sorgen, dass sich die Zellen teilen.
Anschließend werden sie auf geeigneten Gewebeunterlagen ausgestreut und
die Differenzierung ausgelöst. Schließlich muss man dafür sorgen,
dass die Zellen ihre spezialisierte Funktion beibehalten. Für jeden
dieser drei Schritte benötigt man unterschiedliche Materialien.
- Die
Zellteilung findet in herkömmlichen Kulturschalen statt. Man verwendet
kleine Gewebestücke, die ca. 1-2 mm dick sind und kultiviert werden können.
Einzelne Zellen müssen durch Proteasen isoliert werden. Die Zellen
teilen sich am besten in statischem Medium, das Wachstumsfaktoren und
Blutserum enthält. Die Konzentrationen der Elektrolyte entsprechen
weitgehend denen, die in wachsendem Gewebe zu finden sind.
- Die
Expression spezifischer Zellmerkmale erfolgt in vitro meistens unvollständig
oder sogar falsch. Deshalb ist auch hier wichtig, das gewebetypische
Umfeld möglichst genau zu imitieren, um eine optimale Differenzierung
zu erhalten. Die Differenzierung hängt wesentlich von der
Zellverankerung ab. Man benutzt deshalb spezielle Gewebeunterlagen, um
Zellen diese Verankerung zu ermöglichen. Die Unterlagen werden
individuell ausgewählt. Es handelt sich um Filter, Fleece, Metall- oder
Keramikunterlagen, bioabbaubare Materialien oder schwammartige Matrices.
Für dreidimensionale Gewebestrukturen braucht man ein Gerüst aus
Polymeren, das sich nach einigen Wochen oder Monaten zersetzt und vom
spezifischen extrazellulären Gewebe ersetzt wird.[21].
Die Gewebeunterlagen mit den darauf ausgestreuten Zellen werden in
Gewebeträgern befestigt. Diese Gewebeträger verhindern, dass sich das
Gewebe wegen unregelmäßiger Gewebespannung aufrollt[22].
- Weiterhin
muss die richtige Temperatur beachtet werden, die durch eine Wärmeplatte
gewährleistet wird, und natürlich auch die Konzentration der
Elektrolyte. Diese variiert je nach dem, ob sich die Zellen teilen oder
differenzieren sollen. Sie spielt eine besonders wichtige Rolle, da
Elektrolyte mit Rezeptoren in Wechselwirkung treten und somit die
Transkriptionsaktivität beeinflussen können, wie ich bereits
beschrieben habe. Das herkömmliche Kulturmedium IMDM ist für die Zucht
von Gewebe nicht optimal; deshalb verändert man hier die
Elektrolytkonzentration im erforderlichen Maße. Darauf werde ich jedoch
später noch einmal ausführlicher eingehen. Weitere Faktoren, die die
Differenzierung beeinflussen, sind Hormone und Wachstumsfaktoren, die
ebenfalls in optimaler Konzentration vorhanden sein müssen.
- Da
sich im entstehenden Gewebe metabolischer Abfall bilden kann, der das
Gewebe zerstört, wird häufig mit sogenannten Perfusionscontainern
gearbeitet: Die Gewebeträger werden in den Perfusionscontainer gestellt
und können so ständig mit frischem Kulturmedium beliefert werden. Das
verbrauchte Medium, in dem die bereits erwähnten metabolischen und
parakrinen Faktoren wie (α-hydroxid-Buttersäure) enthalten sind,
wird durch Peristaltikpumpen entfernt. Auf diese Weise ist ein ständiger
Mediumsfluss gewährleistet, die gewebetypische Situation wird simuliert
und gleichzeitig wird verhindert, dass das Gewebe durch schädliche
Faktoren zerstört wird.
- Zudem
gibt es spezielle Kulturcontainer, in denen das Gewebe auf beiden Seiten
mit unterschiedlichem Medium behandelt werden kann – auch hier wieder
mit dem Zweck, ein gewebetypisches Umfeld zu schaffen. Man benötigt
Medien, die die Proliferation steuern, ebenso wie Medien, die die
Differenzierung steuern[23].
-
-
- 3.2.2. Welche Art von Zellen benötigt man?
-
- Wenn man
Zellen kultiviert und zur Teilung anregt, beginnen sie sich zu
dedifferenzieren und werden scheinbar reembryonalisiert. Sie werden in
ein jugendliches Stadium zurückversetzt, sind aber nicht totipotent,
d.h., sie können sich auch nicht zu jeder beliebigen Zellart
entwickeln. Deshalb verwendet man Zellen des Gewebetyps, zu dem sich die
gezüchteten Zellen später spezialisieren sollen. Besonders häufig
benutzt man Knorpel-, Knochen- oder Darmzellen, Zellen von Herzklappen
oder Keratinozyten. Bei Menschen, deren Haut bis zu 98 % verbrannt ist,
verwendet man Haut aus Beugefalten, um größere Hautstücke zu züchten,
mit denen die offenen Hautstellen patchworkartig überdeckt werden.
-
-
- 3.3.1. Unterscheidung zwischen Zellteilung
und Zelldifferenzierung
-
- In der Zygote
ist das Erbgut inaktiv, ebenso wie in den ersten Zellen, die aus der
Zygote hervorgehen. Das bedeutet, dass keine Transkription stattfindet,
also auch keine Proteine synthetisiert werden. Transkription findet nur
während der Interphase statt. In dieser Phase liegen die Chromosomen
als lange fädige Strukturen im Zellkern vor. Die RNA-Polymerase findet
überall Startpunkte und kann nur die Gene nicht transkribieren, die
durch Transkriptionsfaktoren, die an Silencer binden, blockiert werden.
- Während der
mitotischen Teilung kondensieren die Chromosomen stark. Durch diese
Kondensation wird die Transkriptionsaktivität eingeschränkt. Es können
keine Proteine mehr synthetisiert werden, da die RNA-Polymerase keine
Gene transkribieren kann.
- Da die
Transkription während der Interphase stattfindet, findet auch die
Zelldifferenzierung während der Interphase statt.
- Wenn durch zu
häufige Teilungen zu wenig Proteine synthetisiert werden, setzt die
Dedifferenzierung ein. Die Zellen verlieren ihre ursprüngliche Funktion
und werden in ein Stadium zurückversetzt, dass sie für die
Reproduktion und damit für den ersten Schritt des Tissue Engineering
hervorragend geeignet macht.
- Die
Differenzierung läuft unter anderen Bedingungen ab, als die
Zellteilung. Die beiden Vorgänge verlaufen nicht parallel. Deshalb muss
jeder einzelne Schritt eingeleitet, koordiniert und beendet werden.
Gerade in der Interphase muss darauf geachtet werden, dass die Faktoren,
die die Entwicklung des Gewebes beeinflussen, möglichst genau mit denen
im lebenden Gewebe übereinstimmen. Denn diese Faktoren treten mit
Rezeptoren in Wechselwirkung und können als Transkriptionsfaktoren an
Silencer, Enhancer oder Promotor binden.
-
-
- 3.3.2. Steuerung der Mitose
-
- Im Körper
findet man in sich entwickelnden Geweben eine sehr hohe Wachstumsaktivität
der Zellen. In adulten Geweben befinden sich dagegen die meisten Zellen
in der Interphase.
Dies ist ein Hinweis dafür, dass die extrazelluläre Matrix für die
Differenzierung eine große Rolle spielt. Sie wird von Zellen produziert
und in ihrer Nachbarschaft ausgeschieden. Es handelt sich dabei um
fadenförmige Strukturen (z.B. Kollagen), die ein Gerüst bilden und
Zellen die Möglichkeit zur Verankerung bieten.
Wenn man die Zellen von ihrer organspezifischen extrazellulären Matrix
isoliert, führt dies wahrscheinlich zur Dedifferenzierung.
Wie bereits erwähnt, werden die Zellen dabei in ein jugendliches
Stadium zurückversetzt und sind deshalb für die Reproduktion
hervorragend geeignet.
- Die
Reproduktion erfolgt in Kulturschalen mit statischem Medium, d.h., es
erfolgt kein ständiger Austausch des Mediums. Das Medium enthält neben
Wachstumsfaktoren auch Serum.
Auf diese Weise kann eine Vervielfältigung garantiert werden. Die
Zellen zeigen eine ausgezeichnete Vitalität mit häufigen mitotischen
Teilungen und bilden innerhalb einiger Tage einen vollständigen
Monolayer. Bisher sind alle herkömmlichen Kulturmedien darauf
ausgerichtet, die Zellproliferation zu unterstützen.
- In einigen Geweben findet eine
Proliferation nur in einem sehr frühen Stadium statt. Im adulten
Stadium verharren die Zellen in einer postmitotischen Phase. Die
Amplifizierung muss deshalb erst ausgelöst werden. Das Kulturmedium
wird an den Mitosezyklus angepasst. Durch Zugabe von Serum (fetal calf
serum) und Wachstumsfaktoren werden die Zellen zur Dedifferenzierung und
zur Teilung angeregt. In herkömmlichem IMDM findet man einen Na+-Wert
von 112 mMol/l, während in Serum 137 mMol/l vorhanden sind. Die
Teilungsrate kann deshalb erhöht werden, wenn man den Na+-Wert
erhöht.
- Na+ wirkt offenbar ebenfalls als Steuerungssubstanz,
da es eine erhöhte Aktivität jener Gene bewirkt, die für die
Zellteilung verantwortlich sind.
-
-
- 3.3.3. Steuerung der Zelldifferenzierung
-
- Sobald man
Zellen in ausreichender Anzahl hat, muss man dafür sorgen, dass sie
spezielle Merkmale ausbilden. Die Differenzierung des Gewebes ist
wichtig, damit bei der späteren Implantation keine Immunabwehr erfolgt
und damit das Gewebe einwandfrei funktioniert. Leider wird die
Differenzierung nicht von einem einzigen löslichen Wachstumsfaktor
beeinflusst, sondern von einer Vielzahl umweltbedingter Stimuli, die
miteinander in Wechselwirkung treten.
- Ein besonders
wichtiger Faktor ist die Zellverankerung. Die Zellen werden nach
ausreichender Teilung auf einer optimalen Unterlage ausgestreut. Die
Unterlagen werden individuell ausgesucht. Die Materialien dafür
variieren von Filtern, Fleecen, bioabbaubaren Polymeren, bis zu schwammähnlichen
Matrices. Für dreidimensionale Gewebestrukturen benötigt man
Fiberfasern. Um Zellen eine noch bessere Verankerung zu ermöglichen, überzieht
man die Gewebeunterlagen häufig mit synthetischen Polymeren, Peptiden
oder Proteinen der extrazellulären Matrix wie Polylysin, Fibronectin,
Chondronectin oder Laminin.
Bei Bedarf können die Gewebeunterlagen in eine bestimmte Form gebracht
werden (wie z.B. in die Form eines Ohres).
- Die
Zellverankerung ist einer der wichtigsten Faktoren, um gute Ergebnisse
in Bezug auf die Merkmalsausbildung zu liefern. In vivo stellt die
extrazelluläre Matrix, die von Zellen abgesondert wird und eine fädige
Struktur aufweist, das Gerüst für die Zellen dar, an das sie sich
anheften können. Die extrazelluläre Matrix ist also jeweils Teil des
spezifischen Umfelds eines Gewebes und muss deshalb als ein wichtiger
Differenzierungsfaktor beachtet werden.
- Weiterhin
muss dafür gesorgt werden, dass die kultivierten Zellen gleichmäßigen
Temperatur- und pH-Werten ausgesetzt sind. Eine Wärmeplatte garantiert
die konstante Temperatur, die meist bei ca. 36°-37°C liegt. Kleinere
Temperaturschwankungen werden von den dicken Wänden des
Kulturcontainers auf ein Minimum reduziert.
- Ein hoher Zusatz an NaHCO3 garantiert im Kohlenstoffdioxidinkubator einen
konstanten pH-Wert von 7.4. Ohne diesen Zusatz würde sich der pH-Wert
außerhalb des Inkubators stark verändern, da im Inkubator 5%
Kohlenstoffdioxid vorhanden sind, im Gegensatz zu 0.3% in der Luft. Außerhalb
des Inkubator muss deshalb die NaHCO3 Menge reduziert werden.
- Die
Gewebeunterlagen mit den darauf ausgestreuten Zellen werden in Gewebeträger
eingespannt. Dabei handelt es sich um sogenannte Halterungsringe und
Spannungsringe mit deren Hilfe die Gewebe eingespannt und befestigt
werden.
-
-
-
- Abb. 5: Gewebeträger:
- Die Gewebeunterlagen mit den darauf ausgestreuten
Zellen werden wie das Fell einer Trommel zwischen zwei Ringen
eingespannt.
-
- Die Träger
werden mit Pinzetten in den Kulturcontainer gestellt. Der Gewebeträger
teilt den Container in eine obere und eine untere Abteilung, so dass das
Gewebe bei Bedarf auf beiden Seiten mit unterschiedlichem Medium
behandelt werden kann, wie es zum Beispiel bei Nierengewebe der Fall
ist.
- Die einzelnen Bestandteile des
Mediums müssen genau auf die Bedürfnisse des Gewebes abgestimmt sein.
So kann zum Beispiel die Expression von Proteinen von Epithelzellen der
Niere beispielsweise stark beeinflusst werden, wenn man zu herkömmlichem
IMDM 12mMol/l NaCl und 17 mmol/l Na-gluconate hinzufügt.
Der Wachstumsfaktor VEGF stimuliert ebenso wie das Hormon Aldosteron
(1x10-7mol/l) oder Vitamin D3 die Gefäßbildung, während
sie in Anwesenheit des Wachstumsfaktors bFGF deutlich gehemmt wird. Auch
Retinsäure trägt wesentlich zur Bildung von Gefäßstrukturen bei.
Trotzdem konnten in der Niere bisher noch keine Rezeptoren für Retinsäure
gefunden werden, ganz im Gegensatz zu Rezeptoren für Vitamin D3 und
Aldosteron.
Das Medium ist Serum-frei.
- In der
Gewebezucht wird mit neu entwickelten Perfusionscontainern gearbeitet,
die im Vergleich zu gewöhnlichen Kultursystemen entscheidende Vorteile
aufweisen. Die Funktionsweise dieser Container ist relativ einfach. Es
handelt sich jeweils um einen Container, der über Silikonschläuche mit
Flaschen verbunden ist. Aus einer Flasche werden mit Hilfe einer
Peristaltikpumpe 0.5-5ml/h Medium in den Container gepumpt, in einer
weiteren Flasche wird das verbrauchte Medium gesammelt. Dadurch wird die
ständige Nährstoffzufuhr gewährleistet, die auch immer optimal ist.
Die typische Situation im lebenden Gewebe kann so sehr viel besser
simuliert werden als in statischen Kulturen. Vor allem aber kann man so
vermeiden, dass toxische Stoffe, die bei der Entstehung des Gewebes
gebildet werden, dem Gewebe schaden oder es sogar völlig zerstören.
Dabei handelt es sich um Metabolite, die freigesetzt werden, wenn künstliche
Gewebeunterlagen abgebaut werden. Das Milieu wird saurer (der pH-Wert
sinkt),
da schädliche Stoffe, wie z.B alpha-hydroxid-Buttersäure, oder weitere
metabolische und parakrine Faktoren freigesetzt werden. Durch den ständigen
Austausch des Mediums werden auch diese Stoffe aus dem Gewebe entfernt.
-
-

-
- Abb. 6:
Perfusionscontainer mit je einer Flasche für frisches und
verbrauchtes Kulturmedium
-
-
- 3.4. Welche Art von Gewebe kann
gezüchtet werden?
-
- Bis jetzt
kann man nur Gewebe züchten, die aus einer einzigen Art von Zellen
bestehen. Die berühmte Haut aus der Tube ist ein gutes Beispiel dafür.
Es handelt sich dabei um eine einfache Schicht aus Keratinozyten, die
weder Haarwurzeln noch Schweißdrüsen aufweist und deshalb in ihrer
Funktion erheblich eingeschränkt ist. Patienten, die mit dieser künstlichen
Haut behandelt wurden, haben weder Wimpern noch Augenbrauen, zudem ist
ihre Körpertemperatur überhöht und die Haut weist Risse auf.
- Außerdem
versucht man Knorpel- und Knochenzellen zu züchten. Bei Versuchen,
Nierengewebe zu züchten, ist es bereits gelungen, kleine gefäßartige
Strukturen zu züchten, allerdings steht man auch hier noch am Anfang.
-
-
- 3.5. Welche Herausforderungen
stellen sich in Zukunft?
-
- Welche
Herausforderungen sich in Zukunft stellen werden, ergibt sich schon aus
dem, was ich bisher geschrieben habe: Man muss versuchen, Gewebe zu
erzeugen, in denen verschiedene Zellarten miteinander kommunizieren,
denn unsere Organe bestehen leider nicht aus einem einzigen Typ von
Zellen, sondern sind komplexe Systeme mit vielerlei Aufgaben, für die
verschiedene Zellen nötig sind. Dieses Hindernis wird wohl nicht so
schnell überwunden werden können und so wird es wohl noch sehr lange
dauern, bis es ein „Ersatzteillager“ für unsere Organe gibt.
-
-
-
- Tissue
Engineering ist wahrscheinlich die Variante der Gewebezucht, die am
ehesten akzeptiert ist und als am wenigsten bedenklich eingestuft wird.
In der Frage nach der Herstellung von möglichst kostengünstigem
Gewebe, das im Körper einwandfrei funktioniert, fallen jedoch auch
immer wieder zwei Stichworte: „Therapeutisches Klonen“ und
„Organzucht aus embryonalen Stammzellen“. Hinter diesen Stichworten
verbergen sich Techniken, die sehr viel umstrittener sind und deshalb
weitaus häufiger diskutiert werden, als Tissue Engineering. In dieser
abschließenden bewertenden Stellungnahme zur Organ- und Gewebezucht möchte
ich aus diesem Grund nicht nur auf das Tissue Engineering eingehen,
sondern auch auf die beiden anderen Techniken.
- Dass
viele Menschen auf das Therapeutische Klonen und die Forschung an
embryonalen Stammzellen ablehnend reagieren, liegt sicherlich zum Teil
daran, dass viele Debatten stark emotionalisiert werden. Das Thema wird
in den Medien aufgebauscht, einerseits ist von einer Chance für die
Medizin die Rede, andererseits von der ethischen Problematik. Kritiker
sprechen davon, dass beide Techniken nicht zu verantworten sind. Es wäre
allerdings zu einfach, sämtliche kritischen Ansichten als Unkenrufe
abzutun, stattdessen muss der Sachverhalt sowohl von der
wissenschaftlichen als auch von der ethisch/gesellschaftlichen Seite her
betrachtet werden.
- Zunächst
einmal zur Definition: Was sind embryonale Stammzellen überhaupt? Es
handelt sich hierbei um Zellen, die im Gegensatz zu Somazellen noch
undifferenziert sind und infolgedessen das Potential besitzen, sich zu
jeder beliebigen Körperzelle zu entwickeln. Man hofft nun, mit diesen
Zellen, Gewebe und vielleicht sogar komplette Organe herstellen zu können.
- Diese Gewebe
sind jedoch problematisch, weil sie Immunreaktionen auslösen können.
Denn auch embryonale Stammzellen sind fremde Zellen, gegen die sich der
Körper wehrt. Diese Abstoßungsreaktion möchte man mit einem Trick
verhindern – mit dem Therapeutischen Klonen. Man möchte versuchen,
embryonale Stammzellen künstlich zu erzeugen – aus den Körperzellen
des Patienten. Wenn also ein Patient ein neues Herz bräuchte, würde
folgendes passieren: Aus einer gewöhnlichen Körperzelle des Patienten
wird der Kern vorsichtig entnommen und in eine Eizelle implantiert, die
ebenfalls zuvor entkernt wurde. Mit Stromstößen wird das Erbgut in
seinen Ursprungszustand zurückversetzt und die Zelle beginnt sich zu
teilen. Nun muss der Embryo, der auf diese Weise entstand „nur“ noch
so behandelt werden, dass er sich nicht zu einem vollständigen Menschen
entwickelt, sondern zu einem einzigen Organ – einem Herzen. Natürlich
ist dies heute noch nicht möglich und die Kritiker dieser Technik
versuchen auch zu verhindern, dass es irgendwann möglich sein wird.
- Ihre
Argumente basieren vor allem auf dem Recht der Embryonen, dass sie als
menschliche Lebewesen nicht getötet und zu Forschungszwecken
missbraucht werden dürfen. Darf man Embryonen tatsächlich „zum
Ersatzgewebe oder Organbildner“
umdeuten? Auf diese Weise könnte die Klonierung verharmlost werden,
indem sie „auf scheinbar weniger problematische Handlungen reduziert
wird“.
Dass dies nicht geschehen darf, darüber sind sich die Kritiker einig.
Wenn wir Embryonen züchten, um embryonale Stammzellen zu erhalten,
machen wir sie uns verfügbar. Dabei müssen wir uns im Klaren sein,
dass wir damit per definitionem einen anderen Menschen
instrumentalisieren und ihn uns verfügbar machen, denn auch ein Embryo,
der noch sehr viel eher einem Zellhaufen ähnelt als einem Menschen,
gilt als menschliches Lebewesen.
Menschliches Leben wird im Reagenzglas so weit abgewertet, dass es einer
„verbrauchenden Wissenschaft zum Opfer“
fallen kann – so weit die Befürchtung gegenüber dem Therapeutischen
Klonen. Aus dieser Problematik ergibt sich zwangsweise die Frage nach
der Menschenwürde. Wann beginnt sie und gilt sie auch für Embryonen,
die künstlich erzeugt wurden, um einen anderen Menschen zu heilen? Wenn
man die künstlich geschaffenen Embryonen als menschliche Lebewesen bzw.
als menschliches Leben bezeichnet, muss man auch Embryonen den Schutz
ihrer Würde zugestehen.
- Aus dieser
Perspektive betrachtet, ist das Therapeutische Klonen völlig
ausgeschlossen. Wir dürfen eben nicht alles tun, was wir theoretisch
tun könnten – aus Ehrfurcht vor dem Leben.
- Diese
Argumente sind zwar überzeugend, trotzdem bin ich noch etwas skeptisch,
was hauptsächlich damit zusammenhängt, dass mir diese Argumente als
von der Wirklichkeit zu weit entfernt erscheinen. Die Diskussion findet
zu selten auf einer festen Basis statt und darunter verstehe ich die
eigene Erfahrung. Denn meiner Ansicht nach kann man seine Meinung über
das Therapeutische Klonen sehr schnell ändern, wenn man selbst in eine
Situation kommt, in der einem das Therapeutische Klonen helfen könnte,
z.B., wenn ein Freund oder Verwandter ein neues Organ benötigt. Wenn
man selbst betroffen ist, denkt man oft anders, als wenn es um fremde
Leute geht, die man nie gesehen hat und nie sehen wird.
- Außerdem
finde ich, dass der Patient, der ein neues Organ benötigt, in der
Diskussion um die Würde und den Wert der Embryonen zurücktritt. Hier
ist es nötig, darüber nachzudenken, was Wert überhaupt bedeutet.
Letztendlich bezeichnet der Begriff „Wert“ etwas Abstraktes, etwas,
das nur in unseren Köpfen existiert. Aus diesem Grund hat jeder
Gegenstand und jedes Lebewesen den Wert, der ihm von uns Menschen
beigemessen wird. Das persönliche Umfeld – die Menschen, mit denen
man täglich umgeht und zu denen man eine bestimmte Beziehung aufgebaut
hat - trägt zusätzlich wesentlich zum persönlichen Wert bei. Ein
Patient erhält seinen Wert also zu einem großen Teil von allen
Menschen, die ihn kennen und die ihm sogar nahestehen. Diesen Menschen
ist es wichtig, dass der Patient nicht an Organversagen stirbt, sondern
geheilt werden kann.
- Ein Embryo
kann noch kein vergleichbares persönliches Umfeld haben, da kein Mensch
etwas über ihn weiß. Er ist noch ein unbeschriebenes Blatt. Trotzdem
ist der Umkehrschluss, dass der Embryo wertlos sei, weil ihm das persönliche
Umfeld fehlt, nicht zulässig. Denn der Embryo hat einen Selbstwert, der
aber nicht durch das persönliche Umfeld, sondern durch das
Embryonenschutzgesetz geschützt wird, das Embryonen in gleicher Weise
bereits menschliche Werte beimisst. Sie haben beide den gleichen
Stellenwert – Patient und Embryo.
- Wenn aber ein
Embryo künstlich erzeugt wird, um einem Patienten zu helfen, scheint
dessen Wert in Diskussionen plötzlich gegenüber dem des Patienten größer
zu sein. Denn aus ethischen Gründen möchte man, dass kein Embryo zu
Forschungszwecken missbraucht wird, während es keine ethisch begründeten
Einwände gibt, dass ein Mensch, dem man mit diesem Embryo vielleicht
helfen könnte, an Organversagen stirbt.
- Hier müsste
man nun entscheiden, welches Leben einen größeren Wert besitzt – das
Leben des Embryos oder das des Patienten. Weil man meiner Ansicht nach
bei der ethischen Diskussion diesen Punkt nicht außer Acht lassen kann
und ich es andererseits für äußerst problematisch halte, hier eine
befriedigende Entscheidung zu treffen, ziehe ich die Diskussion auf
wissenschaftlicher Basis vor.
- Denn auch aus
wissenschaftlicher Sicht gibt es neben den vielen Chancen, die das
Therapeutische Klonen bietet, auch einige eindeutige Risiken, die nicht
von der Hand zu weisen sind und die für mich das Therapeutische Klonen
zu einer verhältnismäßig problematischen Technik machen. Zunächst
einmal werden die Möglichkeiten der embryonalen Stammzellen und des
Therapeutischen Klonens laut Prof. Dr. Will Minuth in den Medien sehr
unseriös dargestellt.
- Man müsse
unbedingt beachten, dass die Befunde zu embryonalen Stammzellen immer
von Mäusen stammen würden; mit Menschen habe man keinerlei
experimentelle Erfahrung.
- Zudem seien
die Risiken, mit denen man sich konfrontiert sehen würde - würde man
tatsächlich embryonale Stammzellen zur Züchtung von Gewebe nutzen - zu
groß. Denn bei embryonalen Stammzellen handelt es sich um körperfremde
Zellen und bei einer Implantation könnte es zu Immunreaktionen kommen.
Außerdem besteht gerade bei embryonalen Stammzellen die Gefahr des
Tumorwachstums.
Ein Patient, der ein Ersatzorgan bekommt, das aus embryonalen
Stammzellen gezüchtet wurde, stirbt dann zwar vielleicht nicht mehr an
Organversagen, dafür aber an Krebs.
- Die Probleme,
die beim Therapeutischen Klonen auftreten, sind ähnlich wie die
Probleme bei embryonalen Stammzellen. Allerdings kann hier die Gefahr
der Immunreaktion praktisch ausgeschlossen werden. Dafür sind jedoch
bis zu 300 Versuche nötig, damit sich eine künstlich aus dem Kern
einer Körperzelle und einer entkernten Eizelle erzeugte Zelle tatsächlich
weiterentwickelt.
- Hier könnte
man befürchten, dass die Zellen, die durch mitotische Teilungen daraus
hervorgehen, nur „zweitklassig“ sind. So tritt beispielsweise beim
Reproduktiven Klonen ein Phänomen auf, das für die Lebenserwartung der
Klone eine Rolle spielen könnte: Je häufiger sich Zellen teilen, desto
kürzer werden die Telomere, die Enden der Chromosomen. Folglich sind bei älteren
Lebewesen, deren Zellen sich bereits häufiger geteilt haben, die
Telomere kürzer als bei jungen Tieren. Bei Dolly konnte man jedoch
beobachten, dass ihre Telomere dieselbe Länge aufwiesen wie diejenigen
ihrer Mutter. Diese Tatsache könnte möglicherweise Dollys Leben um
einige Jahre verkürzen. Beim Therapeutischen Klonen ist die Telomerenlänge
allerdings nicht von Bedeutung. Denn hier ist ja der Empfänger des
Gewebes oder des Organs immer zugleich Spender des Zellkerns. Ein zehnjähriges
Kind wird also keine künstlich erzeugte Leber erhalten, deren Zellen
bereits siebzig Jahre alt sind. Das künstliche Organ hat deshalb keine
kürzere „Lebenserwartung“ als der Patient, der das Organ bekommt.
- Dafür müssen
aber andere Risiken beachtet werden, denn wie bei embryonalen
Stammzellen muss auch hier das Problem der Tumorbildung vermieden
werden. Da es sich um Klonierung handelt, müssen zudem Schäden, die während
der Entwicklung auftreten, befürchtet werden.
Laut Prof. Dr. Minuth ist frühestens in zehn bis zwanzig Jahren mit
Ergebnissen auf diesem Gebiet zu rechnen.
- Aus rein
wissenschaftlicher Sicht weisen sowohl die Nutzung von embryonalen
Stammzellen, als auch das Therapeutische Klonen noch erhebliche Mankos
auf, die es zu eliminieren gilt, wenn man mit Hilfe dieser Verfahren
tatsächlich effektiv und kostengünstig Organe züchten will.
- Die
beschriebenen Probleme treten bei Tissue Engineering nicht auf: Ethische
Bedenken sind – auch laut Werner Bartens – keine vorhanden, da man
nicht künstlich versucht Embryonen zu züchten, und deshalb auch ihre Würde
nicht verletzt. Man benötigt keine 300 Eizellen, bis ein Versuch klappt
und es treten keine Immunreaktionen auf, da man Körperzellen des
Patienten als Ausgangsmaterial verwendet. Zudem hat man bereits
experimentelle Erfahrung mit menschlichen Zellen und es liegen schon
Ergebnisse vor, während beim Therapeutischen Klonen sämtliche
Ergebnisse noch Wunschdenken sind, jedenfalls in Bezug auf menschliches
Gewebe. Trotzdem kann auch diese Technik (noch?) nicht als die Lösung
aller Probleme bezeichnet werden.
- Problematisch
ist hier einmal der Faktor Zeit: Es dauert beispielsweise vier Wochen,
um für Menschen, deren Haut zu 98% verbrannt ist, eine ausreichende
Menge an künstlicher Haut zu züchten. Solange müssen diese Menschen
ohne Haut überleben. Und auch wenn die künstliche Haut erfolgreich
transplantiert werden kann, sehen sich die Betroffenen mit erheblichen
Einschränkungen konfrontiert. Denn die künstliche Haut ist längst
kein gleichwertiger Ersatz für die echte. Zunächst hängt dies mit
kosmetischen Defiziten zusammen. Die Hautstücke werden patchworkartig
zusammengesetzt. Dabei bleiben Narben zurück. Außerdem besteht die
Haut, wie gesagt, nur aus einer einzigen Schicht von Zellen, d.h., es
sind weder Haarwurzeln noch Schweißdrüsen vorhanden. Infolgedessen
werden die Patienten weder Wimpern noch Augenbrauen besitzen, ihre Körpertemperatur
ist ständig erhöht und die Haut ist rissig. Abgesehen von den
Problemen, die die fehlenden Schutzfunktionen mit sich bringen, treten
deshalb für die Betroffenen in erster Linie schwere soziale Probleme
auf, da sie Angst haben, sich in der Öffentlichkeit zu zeigen.
- Unter anderem
aufgrund der Probleme, die sich jeweils aus den verschiedenen Techniken
ergeben, stellt sich die Frage, ob es überhaupt sinnvoll ist, Organe
und Gewebe künstlich herzustellen. Schon die Vorstellung, dass Organe
vom Körper isoliert im Labor entstehen, ist vielen unheimlich und nicht
selten wird diese Vorstellung mit einem Gruselkabinett verglichen. Ein
ernstzunehmendes Argument ist dies natürlich nicht, allerdings tauchen
im Zusammenhang mit der Organzucht immer wieder Fragen nach den
gesellschaftlichen Auswirkungen dieser Technik auf.
- Zum einen drängt
sich die Befürchtung regelrecht auf, dass die Menschheit durch
verbesserte Möglichkeiten im medizinischen Bereich immer vollkommener
wird. Wird in Zukunft niemand mehr krank sein? Heißt es nicht mehr
„Nobody is perfect“, sondern „Everybody is perfect“? Und –
Gegenfrage: Soll es den vollkommen gesunden Menschen überhaupt geben?
- Was diese
Frage angeht, können wir vermutlich beruhigt sein. Wir sind gar nicht
in der schwierigen Situation zu beurteilen, ob es den gesunden Menschen
überhaupt geben soll oder nicht. Denn den gesunden Menschen wird es
niemals geben, da es immer wieder neue Krankheiten geben wird, gegen die
man ankämpfen muss. Wenn es im Mittelalter die Pest war, so ist es
heute AIDS. Die Pest ist besiegt, jedenfalls weitgehend. An AIDS dagegen
erkranken immer mehr Menschen und es gibt noch keine Aussicht auf
Heilung. Allerdings: jeder Mensch möchte gerne gesund sein und bisher
war es immer selbstverständlich, dass sich die Medizin darum bemüht
hat, Medikamente gegen alle möglichen Krankheiten zu erfinden. Deshalb
kann man meiner Meinung nach Menschen mit Organschäden davon nicht
ausschließen. Denn warum sollte ein Mensch, der ohne Penicillin an
einer Grippeinfektion sterben würde, ein größeres Recht auf Leben
haben, als jemand, der ein neues Herz braucht.
- Problematischer
ist die drohende (bzw. bereits vorhandene) Überalterung der
Gesellschaft. Je gesünder die Menschen werden, desto älter werden sie
auch. Es wird also immer mehr alte Menschen geben und da gleichzeitig
weniger Kinder geboren werden, besteht kein Ausgleich mehr. Dem kann ich
nur entgegenhalten, dass Tissue Engineering auch und vor allem jungen
Menschen helfen soll, die an unheilbaren Organschäden leiden. Das
Risiko junger Menschen, durch Unfälle Schäden an Organen
davonzutragen, ist vielleicht sogar größer, als das alter Menschen.
Zudem kann vielen alten Menschen auch mit einer Organtransplantation
nicht geholfen werden, da zu viele Organe beschädigt sind oder der Körper
allgemein zu stark geschwächt ist. Denn man darf nicht vergessen, dass
mit jeder Transplantation auch ein sehr hohes Risiko verbunden ist.
Bereits kleinste Fehler können tödlich sein. Ein Mensch ist eben keine
Maschine, bei dem man jedes Einzelteil beliebig austauschen kann. Wenn
auch die Herstellung dieser „Einzelteile“ irgendwann möglich sein könnte,
der „Einbau“ wird wohl immer schwierig bleiben und er ist bei alten
Menschen noch schwieriger. Je älter die Patienten sind, desto seltener
werden deshalb Transplantationen durchgeführt. Verhindert man aber die
Organzucht generell, verhindert man auch, dass junge Menschen geheilt
werden können und verstärkt damit das Problem der Überalterung noch.
Zudem beginnt das Problem der Überalterung nicht erst mit der
Organzucht, denn dieses Problem ist weitaus älter. Es besteht bereits,
seit man versucht, Menschen mit Medikamenten oder Operationen zu heilen,
um damit ihr Leben zu verlängern. Will man das Problem der Überalterung
lösen, muss man – so drastisch es klingen mag – die medizinische
Versorgung ab einem bestimmten Lebensalter generell verbieten, nicht nur
bei Patienten mit Organversagen. Und dies ist wiederum ethisch nicht
vertret- und verantwortbar.
- Soll man also
Organe züchten? Meiner Ansicht nach schon, denn eine Krankheit hat für
die Patienten immer eine Verminderung des Lebensstandards zufolge. Dies
kann sogar so weit gehen, dass Menschen mit Organschäden völlig von
anderen Menschen abhängig sind, was meiner Meinung nach ebenfalls eine
Verletzung der Menschenwürde bedeutet.
- Man sollte
also Organe nicht züchten, um den vollkommen gesunden Menschen zu
schaffen, sondern um kranken Menschen ihr Leben zu erleichtern.
-
- verwendete
Abkürzungen:
- kb: Kilobasen, VEGF: Vascular Endothelial Growth Factor,
bFGF: basic Fibroblast Growth Factor, IMDM: Iscove`s
modified Dulbecco`s medium, z.B.: zum Beispiel,
-
d.h.: das heißt, bzw.: beziehungsweise
-
-
- ANHANG
-
-
- a) Glossar
- b) Literaturliste
- c) Abbildungsnachweis
- d) weiterführende kommentierte
Bibliographie
-
-
-
-
- Aldosteron
- in der Zona glomerulosa der Nebennierenrinde produziertes
(aus Progesteron über Corticosteron) und auch künstlich
synthetisierbares Hormon bekannter chem. Struktur.
-
- Arginin
- Aminosäure
-
- bFGF
- basic Fibroblast Growth
Factor
-
- cAMP
- zyklisches Adenosin-3`,5`-monophosphat:
nimmt als second-messenger eine zentrale Stellung in der hormonalen
Regulation und im Stoffwechsel ein (Aktivierung von Proteinkinasen, die
ihrerseits durch Phosphorylierung inaktive aktivieren).
-
- cGMP
- zyklisches Guanosinmonophosphat
-
- Cystein
- Aminosäure
-
- Dimere
- chem. Verbindung aus zwei
gleichartigen Molekülen
-
- Domäne
- abgegrenzte Region in Makromolekülen
-
- Elektrolyte
- Substanzen, die in wässriger Lösung
oder in geschmolzener Form der elektrolytischen Dissoziation
unterliegen, d.h. zum Teil in positive Kationen und in negativ geladene
Anionen zerfallen.
-
- extrazelluläre
Matrix
- Die extrazelluläre Matrix wird von Zellen produziert und
in ihrer Nachbarschaft ausgeschieden. Es handelt sich um fadenförmige
Strukturen (z.B. Kollagen), die ein Gerüst bilden. Zellen verankern
sich an diesem Gerüst.
-
- Histidin
- Aminosäure
-
- IMDM
- IMDM = Iscove`s modified Dulbecco`s
medium
- Parameter:
- Ca++:
1.15 mMol/l
- K++:
4.25 mMol/l
- Cl-:
85.1 mMol/l
- Na+: 112.3 mMol/l
-
- Interferon
- von
virusinfizierten Zellen abgegebenes Protein, das andere Zellen schützt,
indem es die Entwicklung von Viren hemmt.
-
- Interphase
- der Zeitabschnitt zwischen zwei Mitosen
im Zellzyklus
-
- Leucin
- Aminosäure
-
- Ligand
- von einem
Membranrezeptor bindungsfähige bzw. von ihm zu bindende Substanz.
-
- Lysin
- Aminosäure
-
- Metabolit
- jede im biologischen Stoffwechsel
auftretende niedrigmolekulare Substanz (von den sog. Biopolymeren nur
deren Vorstufen, Abbau- und Bildungsprodukte).
-
- Mitose
- Zellteilung, im Dienste der Wachstums-
und Zellerneuerungsprozesse.
-
- Monolayer
- Einfachschicht, -lage
-
- Phenylalanin
- Aminosäure
-
- Polymer
- chem. Makromolekül, das sich aus
einheitlichen „monomeren“ Molekülen zusammensetzt.
-
- Protozyten
- Zellen der Prokaryonten
-
- Serum
- Blutserum
- Parameter des Serums eines
neugeborenen Kaninchens:
- Ca++: 1.66
mMol/l
- K+:
6.04 mMol/
- Cl-:
99 mMol/l
- Na+:
137 mMol/l (Vgl: Minuth Publikation)
-
- Thyrosin
- Aminosäure
-
- Transkription
- „Umschreibung“ von DNA in mRNA;
Erbinformation wird auf diese Weise beweglich gemacht und kann den
Zellkern verlassen.
-
- Translation
- Übersetzung von mRNA in Polypeptide
-
- VEGF
- Vascular Endothelial Growth
Factor (Wachstumsfaktor)
-
- Zygote
- befruchtete Eizelle
-
-
-
- b)
Literaturliste
-
-
- ACH,
Johann S. / BRUDERMÜLLER, G. / RUNTENBERG, C. (Hrsg.):
„Hello Dolly ?
- Über das
Klonen“, 1. Aufl., Erstausg.. - Frankfurt am Main : Suhrkamp, 1998. -
250 S.; (dt.) (Edition Suhrkamp ; 2060 : Standpunkte), ISBN
3-518-12060-3
BAYRHUBER, Horst / KULL, Ulrich:
“Linder Biologie”
-
Hannover (Schroedel Verlag GmbH)1998
-
- CALAPRICE,
Alice (Hrsg.): “Einstein sagt – Zitate, Einfälle, Gedanken”, München,
(Piper Verlag
- GmbH), 1997,
272 S., ISBN 3-492-22805-4
-
- CHRIST,
Bodo / WACHTLER, Franz: “Medizinische Embryologie – Molekulargenetik
–
- Morphologie
– Klinik”, Wiesbade (Ullstein Medical Verlagsgesellschaft mbH &
Co.), 1998,
- 348 S.
-
- CZIHAK,
Gerhard / LANGER, Helmut / ZIEGLER, Hubert (Hrsg.):
- “Biologie”
-
Berlin Heidelberg (Springer) 1996
-
- DARNELL,
James / LODISH, Harvey / BALTIMORE, David: “Molekulare
- Zellbiologie”
-
Berlin, New York (de Gruyter) 1993
-
- HAGEMANN,
Rudolf: “Allgemeine Genetik”
-
Heidelberg, Berlin (Spektrum
Akademischer Verlag) 1999
-
- SCHWEGLER,
Johann S.: “Der Mensch – Anatomie und Physiologie. Schritt
- für Schritt
Zusammenhänge verstehen.”
- Stuttgart
(Thieme) 1998
-
- STIEGLER,
Gabor: “Stichwort: Klonen”
-
München (Heyne) 1997
-
- WATSON,
James D. / GILMAN, Michael / WITKOWSKI, Jan / ZOLLER,
- Mark: “Rekombinierte DNA”
- Heidelberg (-Berlin-Oxford)
(SpektrumAkademischer Verlag) 1993
-
-
- Lexikon
Medizin, München – Wien – Baltimore, (Urban &
Schwarzenberg), 1997
-
- Zeitungsartikel
-
- BARTENS,
Werner: Das Schweigen der Lämmer
- in Badische
Zeitung 10.3.2001
-
- Organtransplantation geht neue Wege
- in Süddeutsche
Zeitung 10.2.1999
-
- Frischzellenkur für das Großhirn
- in Süddeutsche
Zeitung 19.10.1999
-
- Haut aus der Retorte
- in Süddeutsche
Zeitung 9.3.1999
-
-
- Internet
-
- ARBEITSGRUPPE
PROF. W. MINUTH: Tissue engineering
-
http://www.biologie.uni-regensburg.de/Anatomie/Minuth/forsch.htm
-
-
-
- KLOTH, Sabine; GERDES, Johannes;
WANKE, Christiane; MINUTH, Will:
Basic
- fibroblast
growth factor is a morphogenic modulator in kidney
- vessel
development
- 1998
by the International Society of Nephrology
-
- KLOTH,
S.; KOBUCH, K.; DOMOKOS, J.; WANKE, C.; MONZER, J.:
- Polar
Application of Test Substances in an Oganotypic Environment and under
Continous Medium Flow: A New Tissue-based Test Concept for a Broad Range
of Applications in Pharmatoxicology
- Accepted
28 October 1999
-
- KLOTH,
Sabine; MONZER, Jan; ECKERT, Elfriede; WANKE, Christiane; KLEIN, Stefan
J.; MINUTH, Will:
Letter to the Editor – Gastric
- Epithelium
under Organotypic Perfusion Culture
- 1998
Society for In Vitro Biology
-
- MINUTH,
Will; SCHUMACHER, Karl; STREHL, Raimund; KLOTH,
- Sabine: Physological and cell biological aspects of
perfusion culture technique employed to generate differentiated tissues
for long term biomaterial testing and tissue engineering
- Received
23 July 1999; accepted 22 December 1999
-
- MINUTH,
Will; SITTINGER, Michael; KLOTH, Sabine: Tissue
- engineering:
generation of differentiated artificial tissues for biomedical
applications
-
Received: 17 April 1997 / Accepted: 27
May 1997
-
- MINUTH;
Will; STEINER, Pat; STREHL, Raimund; SCHUMACHER, Karl; DE
- VRIES,
Uwe; KLOTH, Sabine: Modulation of Cell Differentiation
- in
Perfusion Culture
- Experimental
Nephrology 1999
-
- REISCHL,
Judith; PRELLE, Katja; SCHÖL, Heidrun; NEUMÜLLER,
- Christine;
EINSPANIER, Ralf; SINOWATZ, Fred; WOLF; Eckhard:
Factors affecting proliferation and dedifferentiation of primary bovine
oviduct epithelial cells in vitro
- Received:
25 September 1998 / Accepted: 17 November 1998
-
- ROTTER,
Nicole; AIGNER, J.; NAUMANN, A.; PLANCK, H.; HAMMER,
- C.; BURMESTER, G.; SITTINGER, M.:
Cartilage reconstruction in head and neck surgery: Comparison of
resorbable polymer scaffolds for tissue engineering of human septal
cartilage
- Received
5 August 1997; accepted 17 April 1998
-
- SITTINGER,
M.; SCHULTZ, O.; KEYSZER, G; MINUTH, W.;
- BURMESTER, G: Artificial tissues in perfusion culture
- Wichtig
Editore, 1997
-
-
-
-
- Außerdem berufe ich mich auf e-mails
und Gespräche von und mit Prof. Dr. Will Minuth, Werner Bartens und
Frieda Chen.
-
- Prof. Dr. Minuth leitet am Anatomischen
Institut der Universität Regensburg eine Arbeitsgruppe zu Tissue
Engineering.
- Werner Bartens ist
Wissenschaftsjournalist in Freiburg.
- Frieda Chen war 1999 als Postdoc am
Max-Planck-Institut für Immunbiologie in Freiburg tätig.
-
-
- c)
Abbildungsnachweis
-
- Deckblatt: AOL
- Abb. 1: WATSON, James D. / GILMAN,
Michael / WITKOWSKI, Jan / ZOLLER,
- Mark:
Rekombinierte DNA, Heidelberg (-Berlin-Oxford), (SpektrumAkademischer
Verlag)
- 1993, 596 S., S.148
- Abb. 2:
WATSON, James D. / GILMAN, Michael / WITKOWSKI, Jan / ZOLLER,
- Mark:
Rekombinierte DNA, Heidelberg (-Berlin-Oxford), (SpektrumAkademischer
Verlag)
- 1993, 596 S., S.148
- Abb. 3: WATSON, James D. / GILMAN,
Michael / WITKOWSKI, Jan / ZOLLER,
- Mark:
Rekombinierte DNA, Heidelberg (-Berlin-Oxford), (SpektrumAkademischer
Verlag)
- 1993, 596 S.,
S.148
- Abb. 4:
WATSON, James D. / GILMAN, Michael / WITKOWSKI, Jan / ZOLLER,
- Mark:
Rekombinierte DNA, Heidelberg (-Berlin-Oxford), (SpektrumAkademischer
Verlag)
- 1993, 596 S., S.148
- Anhang: http://www.minucells.de/page07.htm
-
-
-
- d)
weiterführende kommentierte Bibliografie
-
- BUBLATH,
Joachim: Das Geheimnis des Lebens. Genetik
– Urknall – Evolution.
- Wien (Ueberreuter)
1994
- viele Bilder zum Thema
-
- CAVALLI-SFORZA,
Luca und Francesco:
Verschieden und doch gleich. Ein
- Genetiker
entzieht dem Rassismus seine Grundlage.
- München
(Knaur) 1994
- für
mein Thema nicht geeignet
-
- CERNAJ,
Ingeborg und Josef: Am Anfang war Dolly. Geklont und manipuliert –
Leben als
- Spielzeug der
Wissenschaft.
- München
(Wilhelm Heyne Verlag) 1997
- kurze Darstellung der Gentechnik, leider
etwas zu oberflächlich, daher zur Informationssuche nicht sehr geeignet
-
- CZIHAK,
Gerhard / LANGER, Helmut / ZIEGLER, Hubert (Hrsg.):
- Biologie
-
Berlin Heidelberg (Springer) 1996
-
Hintergrundinformationen
über biologische Zusammenhänge
-
-
- GASSEN,
Hans Günter / KEMME, Michael: Gentechnik. Die Wachstums-
- branche der
Zukunft.
- Frankfurt/Main
(Fischer) 1996
- Darstellung
der Gentechnik aus wissenschaftlicher Sicht
- enthält
wenig Informationen über tissue engineering
- für
viele andere Themen gut geeignet
-
- HEBERLE-BORS,
Erwin: Herausforderung Gentechnik
-
Wien (Holzhausen) 1996
- Informationen über Gentherapie, aber
nichts Brauchbares über Tissue Engineering
-
- HINGST,
Wolfgang: Zeitbombe Gentechnik
-
Wien (Orac) 1988
- - absolut ungeeignet, da das Buch fast nur
aus einzelnen Zitaten zusammengesetzt ist
- - keine neutrale Darstellung, viele
Vorurteile, reißerisch geschrieben
-
- KUHN,
Wilfried/ MÜLLER, Thomas: Morbus Parkinson
-
Stuttgart (Georg Thieme Verlag) 1998
- Beschreibung der Parkinsonschen Krankheit
und Behandlungsmöglichkeiten
-
-
- MURKEN,
Jan / CLEVE, Hartwig (Hrsg.): Humangenetik
-
Stuttgart
(Enke) 1988
-
wissenschaftliche
Grundlagen der Humangenetik
-
- PODSCHUN,
Trutz Eyke: Sie nannten sie Dolly. Von Klonen, Genen und
- unserer
Verantwortung.
- Weinheim
(WILEY-VCH)1999
- -
leicht verständlich und differenziert geschrieben
- -
sehr gut geeignet, wenn man sich über Gentechnik informieren will
- -
leider nur wenige Informationen über Gewebszucht
- Das
Buch ist für Laien geschrieben worden, vereinfacht aber nicht zu stark.
-
- DR.
PROWALD, Katja: Gentechnik. Riesentomaten am Nordpol? Erforschung
jahrtausendealter
- Chromosomen
und Heilung von Erbkrankheiten:
- Gentechnik
– Eine Wissenschaft mit vielen Gesichtern.
- München (Südwest
Verlag) 1994
- viele
Bilder zum Thema
-
- SCHWEGLER,
Johann S.: Der Mensch – Anatomie und Physiologie. Schritt
- für Schritt
Zusammenhänge verstehen.
- Stuttgart
(Thieme) 1998
- Anatomie
des Menschen (Entwicklung des Embryos)
-
- STIEGLER,
Gabor: Stichwort: Klonen
-
München (Heyne) 1997
-
kurze Erläuterungen
zur Technik des Klonens
-
knapp und präzise
-
-
- Zeitungsartikel / Zeitschriften
etc. (nach Datum geordnet)
-
- MEICHSNER,
Irene: Herzen, Lebern und Nieren von Schweinen?
- in Badische
Zeitung 11.8.1998
-
- KNÜPFER,
Uwe: Unsterblicher Mr. Seed
- in Badische
Zeitung 8.9.1998
-
- KNÜPFER,
Uwe:
Gott hat mir gesagt, was ich zu tun habe
- in Badische
Zeitung 24.9.1998
-
- SCHULTE,
Markus C.:Erfolge wie bei Verwandten
- in Süddeutsche
Zeitung 3.11.1998
- -
Informationen über die Probleme bei Organtransplantationen/ Übertragung
von Blut-
-
Stammzellen
- -Gründe,
warum es notwendig ist, Gewebe zu transplantieren, dass dem körpereigenen
-
möglichst ähnlich ist
-
- WORMER,
Holger: Embryonen von der Stange
- in Süddeutsche
Zeitung 6.11.1998
- -
Informationen zur Stammzellenzüchtung
- -
differenzierter Artikel, Aspekte der Forschung und Ethik, Einbeziehung
der gesetzlichen
-
Lage
-
- WORMER,
Holger: Embryonenschutz in Gefahr
- in Süddeutsche
Zeitung 6.11.1998
- Kommentar,
keine Hintergrundinformationen
-
- Kopf-an-Kopf-Rennen von Mensch und Tier
- in Süddeutsche
Zeitung 10.11.1998
-
- KOCH,
Klaus: Gene mitten ins Herz
- in Süddeutsche
Zeitung17.11.1998
-
- BERNDT,
Christina: Das zweite Ich aus der Kühltruhe
- in Süddeutsche
Zeitung 24.11.1998
-
- Durchbruch bei der Leukämietherapie
- in Badische
Zeitung 1.12.1998
-
- BERNDT,
Christina: Sicherheit vor Erfolg?
- in Süddeutsche
Zeitung 8.12.1998
-
- Experten: Klonforschung am Menschen
fortsetzen
- in Süddeutsche
Zeitung 9.12.1998
-
- ERMERT,
Monika: Tor zur Embryonenforschung?
- in Süddeutsche
Zeitung 15.12.1998
-
- BAIER,
Tina: Klone fast vom Fließband
- in Süddeutsche
Zeitung 15.12.1998
-
- Muskelkraft aus der Genspritze
- in Süddeutsche
Zeitung 22.12.1998
-
- KOCH,
Klaus: Doch Klone von Menschen?
- in Süddeutsche
Zeitung 29.12.1998
-
- Die Politik der Klonforscher
- in Süddeutsche
Zeitung 21.1.1999
-
- „Dolly“-Schöpfer
Wilmut will menschliche Embryos klonen
- in Süddeutsche
Zeitung 21.1.1999
-
- GOLDHAHN,
Sabine: Blut entsteht aus Hirn
- in Süddeutsche
Zeitung26.1.1999
-
- AIDS-Viren und andere heilsame
Gen-Taxis
- in Süddeutsche
Zeitung 2.2.1999
-
- Organtransplantation geht neue Wege
- in Süddeutsche
Zeitung 10.2.1999
-
- Klonschaf mit Genveränderung
- in Süddeutsche
Zeitung 2.3.1999
-
- Münchner Forscher sind Leukämie auf
der Spur
- in Süddeutsche
Zeitung 4.3.1999
-
- SIEGMUND-SCHULTZE,
Nicola: Die Organtransplantation
- in Süddeutsche
Zeitung 9.3.1999
-
- SIEGMUND-SCHULTZE,
Nicola: Gefährliche Xenotransplantationen
- in Süddeutsche
Zeitung 16.3.1999
-
- WORMER,
Holger: Keine Embryos im Genlabor
- in Süddeutsche
Zeitung 20./21.2.1999
-
- Der eigene Hals hilft bei Parkinson
- in Süddeutsche
Zeitung 25.4.1999
- Hintergrundinformationen
zu Parkinson
-
- Viren im Schwein: Risiko für
Transplantationen
- in Süddeutsche
Zeitung 25.4.1999
-
- Embryozellen im Test gegen Parkinson
- in Süddeutsche
Zeitung 27.4.1999
-
- Erstmals Kälber aus Zellen in Milch
geklont
- in Süddeutsche
Zeitung 27.4.1999
- Klonierung
-
- THURAU,
Martin: Streitfall Embryonenforschung
- in Süddeutsche
Zeitung 5.5.1999
-
- BANTLE,
Oliver (verantw.): Embryonen – ein Rohstoff für die Wissenschaft
- in Süddeutsche
Zeitung 11.5.1999
- ausführlicher,
differenzierter Artikel
- Berücksichtigung
vieler Aspekte
-
- BECK,
Ulrich: Ethik und Genetik
- in Süddeutsche
Zeitung3./4.7.1999
-
- Genspritze gegen Tbc
- in Süddeutsche
Zeitung 10.8.1999
-
- Vom Schwein zum Menschen
- in Süddeutsche
Zeitung 24.8.1999
-
- SCHULTE,
Markus C.: Frischer Geist aus dem Labor
- in Süddeutsche
Zeitung2.9.1999
-
- Frischzellenkur für das Großhirn
- in Süddeutsche
Zeitung 19.10.1999
- Zelldifferenzierung
-
- BARTENS,
Werner: Herz oder Hirn – Gewebe auf Bestellung
- in Badische
Zeitung 24.6.2000
-
- Therapeutisches Klonen im Tierversuch
gelungen
- in Badische
Zeitung 15.8.2000
-
- BARTENS,
Werner: Ersatzgewebe mit Hilfe von Stammzellen
- in Badische
Zeitung 15.8.2000
- Klonierung
-
- BARTENS,
Werner: Revolutionäre Zellen (Dossier)
-
in Die Zeit 24.8.2000
-
sehr ausführlich
-
- Alle
Artikel ohne Kommentar sind entweder zu oberflächlich oder für mein
Thema nicht geeignet.
-
-
- Internet
-
- Arbeitsgruppe
Gelenkknorpelersatz:
Michael Joneleit (Arzt im Praktikum)
Andreas Rücker (wissenschaftlicher Assistent)
PD Dr. Norbert Michael Meenen (Oberarzt Abteilung Unfallchirurgie):
Gelenkknorpelersatz
- http://www.uke.uni-hamburg.de/kliniken/chirurgie/unfall/Forschung/exp_unfall.de.htm#Gelenkknorpelersatz
- Herstellung
von Gelenkknorpel
- der
Artikel ist leider etwas kurz
-
- FRAU
DR. MED. TANCZOS: Abteilung Plastische und Handchirurgie
- http://www.ukl.uni-freiburg.de/chi/pla/plaframe.html
-
Hautherstellung
-
leider etwas kurz
-
Minuth: Tissue Engineering
-
Czihak, Gerhard / Langer, Helmut / Ziegler, Hubert (Hrsg.):
“Biologie” Berlin Heidelberg (Springer) 1996, S.11
-
Vgl.: Bodo Christ, Franz Wachtler, “Medizinische Embryologie –
Molekulargenetik – Morphologie - Klinik”, Wiesbaden, 1998,
Ullstein Medical, 348 Seiten, DM 54, S. 25 f.
-
hrsg. von Prof. Dr. Horst Bayrhuber, Prof. Dr.Ulrich Kull, “Linder
Biologie”, Hannover, 1998 (21. Aufl.), Schroedel Verlag GmbH, S.
342
-
Vgl. Bodo Christ, Franz Wachtler, a.a.o., S. 8f.
-
Vgl. Rudolf Hagemann, “Allgemeine Genetik”,Heidelberg; Berlin,
1999 (4. Aufl.), Spektrum, Akad. Verlag, S.394 f.
-
James Darnell, Harvey Lodish, David Baltimore, “Molekulare
Zellbiologie”, Berlin, 1993, Walter de Gruyter,1277 Seiten, S.465
f.
-
James D. Watson, Michael Gilman, Jan Witkowski, Mark Zoller
“Rekombinierte DNA”, Heidelberg-Berlin-Oxford, 1993 (2.Aufl.),
Spektrum Akademischer Verlag, 596 Seiten, S.147
-
Vgl.: James Darnell, Harvey Lodish, David Baltimore, a.a.o., S.466f.
-
Vgl. James D. Watson, Michael Gilman, Jan Witkowski, Mark Zoller,
a.a.o.; S.149
-
Vgl.: James Darnell, Harvey Lodish, David Baltimore, a.a.o., S.469f.
-
Vgl. James D. Watson, Michael Gilman, Jan Witkowski, Mark Zoller,
a.a.o.; S.307ff.
-
Vgl: Bodo Christ, Franz Wachtler, a.a.o., S.12f.
-
Arbeitsgruppe Prof. Dr. W. Minuth, Forschungstätigkeit, “Tissue
Engineering – Herstellung von künstlichen Geweben für die
Biomedizin”
- Vgl.:
“Frischzellenkur für das Großhirn”, SZ, 19.10.1999
-
Vgl.: Will W. Minuth, Michael Sittinger, Sabine Kloth, “Tissue
Engineering: generation of differentiated tissues for biomedical
applications”, Cell Tissue Res (1998) 291:1-11; © Springer-Verlag
1998, Received: 17 April 1997 / Accepted: 27 May 1997
-
Vgl.: Will W. Minuth, Michael Sittinger, Sabine Kloth, “Tissue
Engineering: generation of differentiated tissues for biomedical
applications”, Cell Tissue Res (1998) 291:1-11; © Springer-Verlag
1998, Received: 17 April 1997 / Accepted: 27 May 1997
-
Vgl.: S. Kloth, K. Kobuch, J. Domokos, C. Wanke, J. Monzer, a.a.o.
-
Vgl.: Will W. Minuth, Michael Sittinger, Sabine Kloth, a.a.o.
-
Vgl.: Telefongespräch mit Prof. Dr. Minuth am 27. März
2001
-
Vgl.: Will W. Minuth, Michael Sittinger, Sabine Kloth, a.a.o.
-
Sabine Kloth, Johannes Gerdes, Christiane Wanke, Will W. Minuth,
“Basic fibroplast growth factor is a morphogenic modulator in
kidney vessel development”, Kidney International, Vol. 53 (1998),
pp.970-978, © 1998 by the International Society of Nephrology
-
Vgl.: Will W. Minuth, Karl Schumacher, Raimund Strehl, Sabine Kloth,
a.a.o.
-
Telefongespräch mit Prof. Dr. Minuth am 27. März 2001
-
Vgl.: Will W. Minuth, Pat Steiner, Raimund Strehl, Karl Schumacher,
Uwe de Vries, Sabine Kloth, “Modulation of Cell Differentiation in
Perfusion Culture”, Exp Nephrol 1999;7:394-406, ©1999S. Karger
AG, Basel
-
Vgl.: Will W. Minuth, Karl Schumacher, Raimund Strehl, Sabine Kloth,
a.a.o.
-
Vgl.: Will W. Minuth, Michael Sittinger, Sabine Kloth, a.a.o.
-
Vgl.: Will W. Minuth, Pat Steiner, Raimund Strehl, Karl Schumacher,
Uwe de Vries, Sabine Kloth, a.a.o.
-
Vgl.: Will W. Minuth, Michael Sittinger, Sabine Kloth, a.a.o.
-
Vgl.: Will W. Minuth, Karl Schumacher, Raimund Strehl, Sabine Kloth,
a.a.o.
-
Vgl.: Will W. Minuth, Michael Sittinger, Sabine Kloth, a.a.o.
-
Vgl.: Sabine Kloth, Johannes Gerdes, Christiane Wanke, Will W.
Minuth, a.a.o.
-
Vgl.: Will W. Minuth, Karl Schumacher, Raimund Strehl, Sabine Kloth,
a.a.o.
-
Vgl.: Telefongespräch mit Prof. Dr. Minuth am 27. März 2001
-
Vgl.: Regine Kollek, “Klonen ist Klonen – oder nicht?”, in
Hello Dolly? Über das Klonen, hrsg. v. J.S. Ach, G. Brudermüller
und C. Runtenberg, (es 2060), Frankfurt 1998, (251 S.);
S.21
-
Regine Kollek, a.a.o., s.22
-
e-mail von Werner Bartens
-
Vgl.: Telefongespräch mit Prof. Dr. Minuth am 27. März 2001
-
Vgl.: Regine Kollek, a.a.o., S.32 f.
-
Vgl.: e-mail von Frieda Chen am 13.10.1999
-
Vgl.: Telefongespräch mit Prof. Dr. Minuth am 27. März 2001 und
“Das Schweigen der Lämmer” (Interview mit Ian Wilmut), BZ,
10.3.2001
-
Vgl.: Telefongespräch mit Prof. Dr. Minuth am 27. März
2001
|
-

|